
STUDIES
SCIENTIFIC & SOCIAL

STUDIES
SCIENTIFIC & SOCIAL
BY
ALFRED RUSSEL WALLACE
LL.D., D.C.L., F.R.S., ETC.
IN TWO VOLUMES.—VOLUME I
WITH NUMEROUS ILLUSTRATIONS
London
MACMILLAN AND CO., LIMITED
NEW YORK: THE MACMILLAN COMPANY
1900
All rights reserved
RICHARD CLAY AND SONS, LIMITED,
LONDON AND BUNGAY.
PREFACE
THE present work consists mainly of reprints of the more important articles I have contributed to reviews and other periodicals during the thirty-five years from 1865 to 1899.
I have ventured to call them "Studies," because the larger part of them deal with problems in which I have been specially interested, and to the comprehension and solution of which I have devoted much time and thought. Many of these problems are connected with the modern theory of evolution, others with important geological and physical theories, others again with educational, political, or social questions. They are dealt with either in the way of exposition or criticism, and in several cases they contain novel views or fresh arguments, strengthening the case in favour of some of the disputed theories.
In order to make the subjects discussed more interesting to the general reader, I have, wherever possible, introduced copious illustrations, and this has led me in many cases so to modify and enlarge the original article as to render it a new piece of work. Those which did not lend themselves to illustration have received a careful revision.
I have now only to thank the various editors and proprietors of periodicals (or books) in which the articles or parts of them first appeared, for permission to reprint them.
I have also to express my thanks to the numerous friends, the scientific societies and the publishers and authors of illustrated works, for the various photographs or engravings I have been permitted to make use of for these volumes. As regard both text and illustrations full acknowledgment is made in the table of contents, and sometimes, where it seemed specially needed, in the body of the work.
A. R. W.
July, 1900.
CONTENTS
| EARTH STUDIES. | ||
| CHAPTER | PAGE | |
| I. | INACCESSIBLE VALLEYS (first appeared in the Nineteenth Century, March, 1893) | 1 |
| II. | THE PERMANENCE OF OCEANIC BASINS (Natural Science, August, 1892) | 28 |
| III. | OUR MOLTEN GLOBE (Fortnightly Review, November, 1892) | 40 |
| IV. | THE ICE AGE AND ITS WORK. ERRATIC BLOCKS AND ICE-SHEETS (Fortnightly Review, November, 1893) | 59 |
| V. | THE ICE AGE AND ITS WORK. GLACIAL EROSION OF LAKEBASINS (Fortnightly Review, December, 1893) | 94 |
| VI. | THE GORGE OF THE AAR AND ITS TEACHINGS (Fortnightly Review, August, 1896) | 129 |
DESCRIPTIVE ZOOLOGY. |
||
| VII. | MONKEYS: THEIR AFFINITIES AND DISTRIBUTION (Contemporary Review, December, 1881) | 146 |
| VIII. | THE DISGUISES OF INSECTS (Science Gossip, September 1, 1867) | 185 |
| PLANT DISTRIBUTION. | ||
| IX. | ENGLISH AND AMERICAN FLOWERS (Fortnightly Review, October, 1891) | 199 |
| X. | FLOWERS AND FORESTS OF THE FAR WEST (Fortnightly Review, December, 1891) | 213 |
ANIMAL DISTRIBUTION. |
||
| XI. | THE PaLæARCTIC AND NEARCTIC REGIONS (Natural Science, June, 1894) | 235 |
| XII. | THE BEETLES OF MADEIRA AND THEIR TEACHINGS (Transactions of the Entomological Society of London, 1871) | 250 |
| XIII. | EVOLUTION AND THE DISTRIBUTION OF ANIMALS (Nineteenth Century, February, 1879) | 265 |
THEORY OF EVOLUTION. |
||
| XIV. | THE ORIGIN OF SPECIES AND GENERA (Nineteenth Century, January, 1880) | 283 |
| XV. | A CRITIC OF NATURAL SELECTION ANSWERED (Natural Science, September, 1894) | 305 |
| XVI. | ARE ACQUIRED CHARACTERS INHERITED? (Fortnightly Review, April and May, 1893) | 315 |
| XVII. | THE METHOD OF ORGANIC EVOLUTION (Fortnightly Review, February and March, 1895) | 345 |
| XVIII. | THE PROBLEM OF UTILITY: ARE SPECIFIC CHARACTERS ALWAYS OR GENERALLY USEFUL? (Journal of the Linnean Society, 1896) | 378 |
| ANTHROPOLOGY. | ||
| XIX. | THE POLYNESIANS AND THEIR MIGRATIONS (Quarterly Journal of Science, April, 1867, much enlarged). | 399 |
| XX. | NEW GUINEA AND ITS INHABITANTS (Contemporary Review, February, 1879, much enlarged) | 416 |
| XXI. | AFFINITIES AND ORIGIN OF THE AUSTRALIAN AND POLYNESIAN RACES (Australasia, Vol. 1, Chap. v. section 10 (with large additions) | 461 |
| SPECIAL PROBLEMS. | ||
| XXII. | THE PROBLEM OF INSTINCT (Natural Science, March, 1897) | 497 |
| XXIII. | HUMAN SELECTION (Fortnightly Review, September, 1890) | 509 |
ERRATA.
VOL. I
LIST OF ILLUSTRATIONS.
| No. 23, | for | "Swainy" read | "Swamy." |
| 57, | „ | "Geological" read | "Geographical." |
| 58, | „ | do. | do. |
LIST OF ILLUSTRATIONS
IN VOL. I
| FIG | PAGE | |
| 1. | THE WATER-SLIDE, DOONE VALLEY Facing | 2 |
| 2. | EL CAPITAN, YOSEMITE VALLEY | 6 |
| 3. | SENTINEL ROCK, YOSEMITE VALLEY | 8 |
| 4. | VIRGIN'S TEARS WATERFALL, YOSEMITE VALLEY | 9 |
| 5. | GOVETT'S LEAP, N.S. WALES Facing | 12 |
| 6. | VIEW FROM GOVETT'S LEAP (From Admiral Maclear) | 15 |
| 7. | THE NORTH DOME, YOSEMITE VALLEY Facing | 22 |
| 8. | DIAGRAM OF HEIGHT OF LAND AND DEPTH OF OCEAN | 32 |
| 9. | GREAT TERMINAL MORAINE IN MASSACHUSETTS (From U.S. Geological Survey) | 63 |
| 10. | MORAINES AT EASEDALE TARN | 64 |
| 11. | ROCHES MOUTONNÉES IN IDAHO (From U.S. Geological Survey) | 66 |
| 12. | STRIATED ROCK-SURFACE AT WASDALE CRAG (From a Photograph by Mr. Godfrey Bingley) | 67 |
| 13. | ROCK GROOVINGS AT BARMOUTH (From a Photograph by Mr. Pecy F.KENDALL) | 70 |
| 14. | ROCK GROOVINGS NEAR BARMOUTH (From an unknown friend) | 71 |
| 15. | MORAINE AT SPECTON (From Photograph by Mr. Kendall) | 83 |
| 16. | BOULDER CLAY ON CHALK, FLAMBOROUGH HEAD (From a Photograph by Mr. Bingley) | 84 |
| 17. | BOULDER CLAY ON UPPER OOLITE, YORKSHIRE (From Photograph by Mr. Bingley) | 85 |
| 18. | A SCANDINAVIAN BOULDER FOUND IN YORKSHIRE (From Photograph by Mr. Kendall) | 86 |
| 19. | SILURIAN ERRATIC ON LIMESTONE, INGLEBOROUGH (From Photograph by Mr. Kendall) | 87 |
| 20. | PERCHED BLOCK IN THE PASS OF LLANBERIS (From Mr. Kendall) | 88 |
| 21. | PERCHED BLOCK NEAR MONTHEY BAS VALAIS (From Falsan's La Periode Glaciaire) | 89 |
| 22. | PERCHED BLOCK NEAR CHAMBERY (From same work) | 89 |
| SECTION OF LAKE COMO (true scale) | 103 | |
| DIAGRAM OF LAKE-FORMS Facing | 116 | |
| MAP OF THE ANCIENT RHONE GLACIER Facing | 128 | |
| 23. | THE AARSCHLUCHT (From Photograph by Mr. Coomara Swainy) | 132 |
| 24. | TERMINAL GORGE OF THE GORNER GLACIER (From Photograph by Mr. Kendall) Facing | 136 |
| 25. | THE GORILLA (From Flower and Lyddeker's Mammals Living and Extinct) | 153 |
| 26. | CHIMPANZEE (From the same work) | 154 |
| 27. | PLUTO MONKEY (From List of Animals in the Zoological Societies' Gardens) | 155 |
| 28 | YELLOW BABOON (From Flower and Lydekker's Mammals Living and Extinct) | 156 |
| 29 | OLD MALE ORANG-UTAN (From Huxley's Man's Place in Nature) | 160 |
| 30 | GIBBON (From Flower and Lydekker's Mammals Living and Extinct) | 162 |
| 31 | Semnopithecus melalophos (From the English Cyclopœdia) | 164 |
| 32 | Macacus leoninus (From List of Animals in the Gardens of the Zoological Society) | 165 |
| 33 | SKULL OF HOWLER (From Flower and Lydekker's work) | 166 |
| 34 | BLACK-HANDED SPIDER MONKEY (From the same work) | 168 |
| 35 | HUMBOLDT'S WOOLLY MONKEY (From the same work) | 169 |
| 36 | WHITE CHEEKED SAPAJOU (From the same work) | 170 |
| 37 | HAIRY SAKI (From Bates' Travels on the Amazon) Facing | 173 |
| 38 | BALD UACARI | |
| 39 | LEMURINE NIGHT MONKEY (From Flower and Lydekker's Work) | 173 |
| 40 | GOLDEN MARMOSET (From the same work) | 175 |
| 41 | GRAY LORIS (From the same work) | 177 |
| 42 | AYE-AYE, SKULL OF (From the same work) | 178 |
| 43 | RING-TAILED LEMUR (From the same work) | 179 |
| 44 | LAPPET MOTH (From Hardwicke's Science Gossip) | 187 |
| 45 | BUFF-TIP MOTH (From same work) | 188 |
| 46 | ORANGE-TIP BUTTERFLY (From same work) | 190 |
| 47 | LEAF BUTTERFLY (From my Malay Archipelago) | 192 |
| 48 | STICK-INSECT (From Science Gossip) | 196 |
| 49 | THE MOTHER OF THE FOREST. Facing | 228 |
| 50 | REDWOOD TREE Facing | 232 |
| 51 | OUTLINE MAP OF THE AZORES | 253 |
| 52 | MAN, TAHITI (Challenger Photograph. By permission) | 406 |
| 53 | GIRL, TAHITI (Challenger Photograph. By permission) | 407 |
| 54 | MAN, HAWAII (Challenger Photograph. By permission) | 408 |
| 55 | GIRL, HAWAII (Challenger Photograph. By permission) | 409 |
| 56 | NATIVE OF TONGA ISLANDS (From Photograph by Mr J. J. Lister) | 411 |
| 57 | MAN OF SAMOA (From Royal Geological Society) | 413 |
| 58 | WOMAN OF SAMOA (From Royal Geological Society) | 414 |
| 59 | Cuscus orientalis (From the Zoological Society of London) | 420 |
| 60 | Dactylopsila trivirgata (From Semon's Australian Bush) | 421 |
| 61 | Dystœchurus pinnatus (From Semon's Australian Bush) | 422 |
| 62 | TREE KANGAROO AND NEW GUINEA BIRDS (From my "Geographical Distribution of Animals") Facing | 425 |
| 63 | KING CHARLES' BIRD OF PARADISE (From Dr. A. B. Meyer, of Dresden) | 427 |
| 64 | FERN-BEARING BIRD OF PARADISE (From Dr. A. B. Meyer, of Dresden) | 428 |
| 65 | ROTHSCHILD'S BIRD OF PARADISE (From Sharpe's Monograph) | 429 |
| 66 | PRINCE RUDOLPH'S BIRD OF PARADISE (From Sharpe's Monograph) | 431 |
| 67 | NATIVE OF DUTCH NEW GUINEA (From Stanford's Australasia) | 439 |
| 68 | PAPUANS, S.W. NEW GUINEA (by Admiral Maclear) Facing | 448 |
| 69 | PAPUAN OF NEW HEBRIDES | 456 |
| 70 | NATIVE OF API (From Challenger Photographs) | 457 |
| 71 | NATIVES OF TIMOR (From a Photograph) | 458 |
| 72 | AUSTRALIAN, NORTHERN TERRITORY (From the Anthropological Institute) | 462 |
| 73 | WOMAN, QUEENSLAND (From the Anthropological Institute) | 463 |
| 74 | AUSTRALIAN HAIRY MAN (Professor Spencer's Photograph) | 466 |
| 75 | HAIRY AINO (From Mr. Hitchcock's Ainos of Japan By permission of the Smithsonian Institution) | 467 |
| 76 | FAMILY GROUP OF AUSTRALIANS (Professor Spencer) Facing | 475 |
| 77 | RUIN OF CAMBODIAN TEMPLE (From Ferguson's Architecture) | 476 |
| 78 | TEMPLE OF BAIOÑ, CAMBODIA (reduced from Garnier's Atlas) Facing | 478 |
| 79 | MODERN CAMBODIANS (From Moura's Royaume de Cambodia) | 482 |
| 80 | SCULPTURES FROM OLD TEMPLES (From Moura's Royaume de Cambodia) | 483 |
| 81 | AINO MAN, URAP (From Hitchcock's Ainos of Japan) | 485 |
| 82 | A VEDDAH SHOOTING (From a Photograph) | 486 |
| 83 | A GROUP OF VEDDAHS (From a Photograph) | 487 |
| 84 | A VEDDAH HUNTER (From a Photograph) | 488 |
| 85 | MAN OF THE ARUNTA TRIBE (Professor Spencer) | 491 |
| 86 | OLD MEN WITH CHURINGA (Professor Spencer) | 492 |
| 87 | NORTH AUSTRALIAN NATIVE (From Admiral Maclear) | 493 |
| 88 | WOMAN AND CHILD, ARUNTA TRIBE (Professor Spencer) | 494 |
| 89 | YOUNG WOMAN, ARUNTA TRIBE (Professor Spencer) | 495 |
STUDIES, SCIENTIFIC AND SOCIAL
CHAPTER I
INACCESSIBLE VALLEYS—A STUDY IN PHYSICAL
GEOGRAPHY
MOST readers of that delightful story Lorna Doone must have been interested in the curious valley occupied by the Doone outlaws as an almost impregnable stronghold. It is described as being about a mile long and a quarter of a mile wide, the nearly-level bottom, through which ran a mountain stream, being bounded on each side by a wall of rock, eighty or a hundred feet high. At the two extremities, these walls approached each other, forming narrow ravines, through which the little river entered and escaped from the valley. At the lower end there was a considerable fall or cataract, over a long steep slope of rock bounded on each side by vertical cliffs, so that the only entrance was up the steep and slippery rocks forming the bed of the torrent, quite impracticable except to a good barefooted climber. At the upper end there appears to have been also some natural barrier, the stream being described as running for a short distance underground; but rude rock-arches had been built over it, forming a kind of tunnel entrance to the valley, which could be easily guarded or blocked up altogether.
If this description applied to any real locality we should have, on a small scale, all the features which characterise
VOL. I.
an "inaccessible valley," the sides formed by rocky precipices, while at the upper and lower ends are narrow gorges rendered impracticable, either by waterfalls, or by the stream filling up the channel at its narrowest portion where the vertical side walls leave no foothold. On a recent visit I found that although the whole surrounding country is very accurately described, the rivers and villages bearing their true names so that the Doone valley can be easily localised, yet it has none of the peculiar features given to it in the story. On a lateral stream that joins the larger valley there is a very charming little cascade which is said to have given the idea of the water-slide up which John Rigg clambered when he first entered the Doone valley. It is well shown in the accompanying photographic print, but in place of the lateral vertical cliffs there are only somewhat rugged slopes very easily ascended, and the little valley above it is open, with gentle slopes on either side and not a cliff to be seen. Neither is there anything corresponding to the deep black pool at the bottom of the cascade, so that the enthusiastic tourist is greatly disappointed, and the people at the nearest house in the valley are said to have been almost driven wild by the constant stream of inquirers till, as a measure of self-defence, they provided refreshments, and in this way obtained some reward for their trouble and loss of time.
Persons who know Exmoor thoroughly say that there is no such valley in any part of the district, and that the talented author has, in this portion of his work, drawn wholly on his imagination for his facts. Nor, so far as I am aware, has such a valley been described in any part of the British Isles, or even in that land of rock-girt valleys and narrow gorges, Switzerland. In fact, considering how very common are each of the four elements required to form an inaccessible valley, it is remarkable how few such valleys exist in any part of the world. These elements are, either a waterfall or a water-blocked gorge at each end, and both sides to be walled by a continuous line of precipices. Valleys with rocky walls on one side and a narrow gorge for outlet are frequent, but then the opposite side has slopes which render it easily accessible. Not unfrequently

FIG. 1.—"THE WATER SLIDE." DOONE VALLEY, NORTH DEVON.
B 2
there is a ravine with waterfalls as the upper outlet also, but in almost every case there is some break in the rock walls on one side or the other with easy slopes for the entrance of men and animals.
The only considerable valleys that can be classed as originally inaccessible—though of course no valley, any more than any mountain, is absolutely so—seem to be, the Yosemite in California, and the valleys of the Grose and Cox rivers in New South Wales. It may, therefore, be interesting to describe these valleys, which are in many ways very remarkable. The theories that have been suggested to account for them may then be considered; and we shall thus be led to discuss the general theory of valley-formation and the peculiar combination of conditions which in these two very dissimilar cases have led to a somewhat similar result.
The Yosemite Valley and its supposed Origin.
The Yosemite valley is a portion of the upper course of the Merced River, which rises near the summit of the Sierra Nevada about 170 miles almost due east of San Francisco. This great mountain range, forming the western edge of the lofty table-land of which the Rocky Mountains form the eastern border, has a very gradual upward slope from the central valley of California, the distance from the foothills to the summit varying from sixty to eighty miles, while the height is from 8,000 to nearly 15,000 feet. This average slope of from 100 to 250 feet in a mile is rendered exceedingly irregular by numerous large winding valleys, some with easy slopes, some more precipitous, and all more or less covered with forest so as to render the journey from one point to another both circuitous and difficult. The higher portion of the Sierra Nevada is usually of granitic rock, lower down are metamorphic slates, followed by enormous beds of late tertiary gravels, which are often covered with great sheets of lava and ashes, bearing witness to the numerous volcanoes on the summit of the range at a period geologically very recent. The Yosemite valley is situated a little
above the middle of the slope, and entirely in the granite region, which is here very wide. It is about seven miles long and from half a mile to a mile wide, the bottom nearly level but rising slightly to the base of the cliffs on either side. These precipices are among the grandest in the world, some of them absolutely perpendicular from base to summit, others with alternate slopes and rock-cliffs, but everywhere equally inaccessible to the ordinary traveller, except in a few places by narrow shelves and steep gullies originally discovered by the Indians and since made into practicable paths or roads. At the lower end the valley becomes narrowed into a deep ravine or cañon for a considerable distance, while at the upper end it branches out into three equally rock-walled valleys with grand waterfalls, leading up to the crest of the mountain range.
This remarkable valley may be said to average about half a mile in vertical depth, but some of the precipices that give it so impressive a character are considerably more than this height, El Capitan at the lower end of the valley being a smooth vertical wall of granite 3,300 feet high with no visible crack or ledge upon it from top to bottom, as shown in the accompanying illustration. Cathedral rock, nearly opposite El Capitan, is 2,600 feet; the Sentinel rock, nearly the middle of the south side of the valley, is over 3,000 feet; while the Half Dome at the upper end of the valley is no less than 4,737 feet high, the upper 1,500 feet of which is quite vertical, while the lower part slopes at an angle of 60° or 70°, and is partly concealed by fallen fragments. The great dome-shaped masses of granite are a characteristic feature of the Sierra Nevada, as they are of some other granitic regions. Nearly opposite the Half Dome is the North Dome, 3,568 feet high, its summit beautifully rounded, but broken lower down so as to show the concentric layers of which it is formed. The Sentinel Dome on the south side is of similar character. The Half Dome is exactly like the other domes in character, but appears as if cut off vertically, leaving the southern half quite perfect and of a fine spherical contour.
The two illustrations (Figs. 3, 4) which face each other over leaf, well serve to show the character of the middle

FIG. 2.—EL CAPITAN, 3,300 FEET. LOOKING DOWN VALLEY.
portion of this wonderful valley, both as to its precipices, rock-pinnacles, and waterfalls; while the numerous slender pines and firs, from 200 to 250 feet high, exhibit the most
characteristic type of the forest vegetation of the Sierra Nevada range.
Professor J. D. Whitney, formerly State Geologist of California, thus characterises the valley in his Yosemite Guide Book:
The principal features of the Yosemite, and those by which it is distinguished from all other known valleys, are: first, the near approach to verticality of its walls; second, their great height, not only absolutely, but as compared with the width of the valley itself; and, finally, the very small amount of talus or débris at the base of these gigantic cliffs. These are the great characteristic features of the Yosemite throughout its whole length; but, besides these, there are many other striking peculiarities and features, both of sublimity and beauty, which can hardly be surpassed, if equalled, by those of any mountain valleys in the world. Either the domes, or the waterfalls of the Yosemite, or any single one of them even, would be sufficient in any European country to attract travellers from far and wide.
The origin of this wonderful valley has been a puzzle even to geologists. After describing the formation of most of the valleys of the Sierra Nevada as being due to denudation, Professor Whitney says:
The eroded cañons of the Sierra, however, whose formation is due to the action of water, never have vertical walls, nor do their sides present the peculiar angular forms which are seen in the Yosemite, as, for instance, in El Capitan, where two perpendicular surfaces of smooth granite, more than 3,000 feet high, meet each other at a right angle. These squarely-cut, re-entering angles, like those below El Capitan, and between Cathedral Rock and the Sentinel, or in the Illilouette cañon, were never produced by ordinary erosion. Much less could any such cause be called in to account for the peculiar formation of the Half Dome, the vertical portion of which is all above the ordinary level of the walls of the valley, rising 2,000 feet, in sublime isolation, above any point which could have been reached by denuding agencies, even supposing the current of water to have filled the whole valley.
He then goes on to discuss the possible agency of ice, which he dismisses as quite inadequate. Valleys formed by fissures of the earth's crust are then discussed, and it is shown that the Yosemite cannot have been formed in this way, partly because it is too wide, and also because there is no correspondence between the opposite sides.
In default of any of the usually accepted theories of
valley-formation, Professor Whitney has been led to adopt one which has hardly yet been recognised by geologists

FIG. 3—SENTINEL ROCK, 3,069 FEET. LOOKING UP VALLEY.
as probable or even possible, and which he describes as follows:
We conceive that, during the process of upheaval of the Sierra, or, possibly, at some time after that had taken place, there was at the Yosemite a subsidence of a limited area, marked by lines of fault
or fissures crossing each other nearly at right angles. In other and more simple words, the bottom of the valley sank down to an unknown depth, owing to its support being withdrawn from underneath during

FIG. 4.—VIRGIN'S TEARS WATERFALL, 3,000 FEET. YOSEMITE VALLEY.
some of those convulsive movements which must have attended the upheaval of so extensive and elevated a chain, no matter how slow we imagine the process to have been.
After showing that subsidence is a well-ascertained fact,
the only difficulty in this place being the great vertical displacement of such a small area, he adds:
By the adoption of the subsidence theory for the formation of the Yosemite we are able to get over one difficulty which appears insurmountable with any other. This is, the very small amount of débris at the base of the cliffs, and even, at a few points, its entire absence. We see that fragments of rock are loosened by rain, frost and other natural causes, along the walls, and probably not a winter elapses that some great mass of detritus does not come thundering down from above, adding no inconsiderable amount to the talus. Several of these great rock-avalanches have taken place since the valley was inhabited. One, which fell near Cathedral Rock, is said to have shaken the valley like an earthquake. This abrasion of the edges of the Valley has unquestionably been going on during a vast period of time; what has become of the detrital material? Some masses of granite now lying in the valley are as large as houses. Such masses as these could never have been removed from the valley by currents of water.…It appears to us that there is no way of disposing of the vast mass of detritus, which must have fallen from the walls of the Yosemite since the formation of the valley, except by assuming that it has gone down to fill the abyss which was opened by the subsidence which our theory supposes to have taken place.
This extraordinary theory, put forth by an experienced geologist in 1874, will probably not be accepted now; but it serves to show that the Yosemite has always been considered a remarkable and exceptional valley which could only have been produced by some equally exceptional causes. A visit to the valley a few years since satisfied the present writer that the modern and now generally accepted theory of valley-formation is quite sufficient to account for the Yosemite, though its features have been rendered almost unique by the peculiar character of the rocks out of which it has been hollowed, combined with the meteorological and physical conditions of the locality, both now and during the latter part of the tertiary epoch. After having described the Australian valleys referred to at the commencement of this chapter, an attempt will be made to show that both are true valleys of denudation.
The Cox and Grose Valleys.
In some respects the valleys carved out of the great sandstone plateau of New South Wales are even more
remarkable than the Yosemite itself. This plateau forms the eastern side of the Blue Mountains, and at its eastern margin is about 1,000 feet above the sea level; but going westward it rises about 100 feet in a mile, so that at its further side, at a distance of twenty-five miles it is 3,400 feet above the sea. This slightly undulating monotonous surface is, however, deeply intersected by widely branching ravines which increase in depth as we proceed westward, and which everywhere present perpendicular crags and cliffs of a very remarkable character. The ravines which discharge their waters into the little river Cox occupy an area of 1,212 square miles. The whole forms the basin of this mountain stream, and is bounded by cliffs increasing from about 1,000 feet near its outlet to about 2,500 feet near its western limits, the valley bottom being not much above the sea level, and the only outlet being through a gorge about a third of a mile wide.
Further to the north is the smaller valley of the Grose, whose diverging ravines interlock, as it were, with those of the Cox, forming a great obstacle to the early explorers in their attempts to cross the plateau. The Grose valley has still grander precipices than that of the Cox, rising at the upper end to 3,000 feet in vertical height. The best account of these valleys is that given in Darwin's work on Volcanic Islands, the last chapter of which is devoted to Australia and other places visited on the homeward voyage. He says:
It is not easy to conceive a more magnificent spectacle than is presented to a person walking on the summit-plains, when without any notice he arrives at the brink of one of these cliffs, which are so perpendicular that he can strike with a stone (as I have tried) the trees growing at a depth of 1,500 feet below him; on both hands he sees headland beyond headland of the receding line of cliff, and on the opposite side of the valley, often at a distance of several miles, he beholds another line, rising up to the same height with that on which he stands, and formed of the same horizontal strata of pale sandstone. The bottoms of these valleys are moderately level, and the fall of the rivers flowing in them, according to Sir T. Mitchell, very gentle. The main valleys often send into the platform great bay-like arms, which expand at their upper ends; and, on the other hand, the platform often sends promontories into the valleys, and even leaves in them great, almost insulated, masses.
So continuous are the bounding lines of cliff, that to descend into some of these valleys it is necessary to go round twenty miles; and into others the surveyors have only lately penetrated, and the colonists have not yet been able to drive in their cattle. But the most remarkable point of structure of these valleys is that, although several miles wide at their upper parts, they generally contract towards their mouths to such a degree as to become impassable. The Surveyor-General, Sir T. Mitchell, in vain endeavoured, first on foot and then by crawling between the great fallen fragments of sandstone, to ascend through the gorge by which the river Grose joins the Nepean; yet the valley of the Grose, in its upper part, as I saw, forms a magnificent basin some miles across, and is on all sides surrounded by cliffs, the summits of which are nowhere less than 3,000 feet above the sea-level. When cattle are driven into the valley of the Wolgan by a path partly cut by the colonists, they cannot escape; for this valley is in every other part surrounded by perpendicular cliffs, and eight miles lower down it contracts from an average width of half a mile to a mere chasm, impassable to man or beast.1
The view here described by Darwin is that from "Govett's Leap" into the Grose Valley, and is shown in the accompanying reproductions from photographs (Figs. 5, 6), but I have not been able to obtain any really good views of the interior of these remarkable cliff-bound valleys. They are also described in Darwin's Naturalist's Voyage, p. 438.
The origin of these valleys appears to have been as great a puzzle to the early explorers as was that of the Yosemite. Sir Thomas Mitchell estimates that 134 cubic miles of rock must have been removed from the valley of the Grose alone; and he remarks on the absence of indication of the agency by which these vast masses of stone have been carried away, there being no accumulations of sand, though there are many huge blocks of rock, scarcely worn by attrition, in the bed of the stream, while in the valleys below, instead of sandy deposits, there is a rich alluvium. Even Darwin was staggered at the idea of these enclosed valleys being hollowed out by aqueous erosion. Neither does he accept subsidence, on account of the numerous irregularly branching arms. The resemblance of the cliffs to those of a bold sea coast suggests marine action, "but
1 The Wolgan is a north-western tributary of the Hawkesbury River, while the Nepean and Grose rivers flow into it from the south-west.

FIG. 5.—GOVETT'S LEAP, BLUE MOUNTAINS, N. S. WALES.
then," he remarks, "occurs the startling difficulty, why has the sea worn out these great, though circumscribed, depressions on a wide platform, and left mere gorges, through which the whole vast amount of triturated matter must have been carried away?" Finally, he suggests, that marine currents often form banks of most irregular form, and so steep that a small amount of subsequent erosion during elevation might form them into cliffs. We must consider, however, that this plateau has certainly been elevated since the latter part of the secondary period, leaving ample time for any amount of denudation; and Mr. Beete-Jukes, in his Sketch of the Physical Structure of Australia, informs us that similar valleys abound throughout the great sandstone formation, both at high and low levels; and they have so exactly the character, in the distribution of their diverging branches, of ordinary streams carrying off the drainage of a slightly inclined surface, that no exceptional origin for them seems needful. This will be more clear when we have discussed the modern theory of valley-formation and the special characteristics of the rocks in which these remarkable valleys have been excavated.
How Valleys are Formed.
One of the most common ideas, when a person sees a deep gorge or ravine bounded by lofty precipices, is, that the rocks have been torn asunder by some earthquake or other subterranean movement. A "convulsion of nature" is almost always referred to in popular descriptions of such scenes. Till recent years even geologists considered that many valleys were so formed. The article on the "Geology of the Alps," by M. Desor, in Ball's Alpine Guide, published in 1870, gives "valleys of disruption" as one of the forms of Alpine valleys, and cites the defile of the Via Mala on the Hinter Rhein, and the valley of the Rhone between Bex and Martigny, as examples. He defines them as "evidently produced by rents that have torn asunder ranges once continuous." Professor Whitney, also, in his Yosemite Guide-Book, speaks of rents or fissures as one of the recognised modes of valley-formation.
Now, however, it is held by most, if not all, geologists that valleys are never formed in this way. It is to the

FIG. 6.—VIEW FROM GOVETT'S LEAP.
late J. Beete-Jukes, Director of the Geological Survey of Ireland, that we owe the full establishment of the principle
that "valleys of all kinds, from the most open to the most narrow and profound, are hollows worn by erosion."1 He was struck by the fact of many of the rivers of the south of Ireland, after running for miles over low plains open to the sea, suddenly turning at right angles, cutting through the hills by deep narrow ravines, and so reaching the sea beyond them. Sometimes even the hills the river cut through were isolated, so that the river might, apparently, have passed round them in either direction. The explanation usually offered of these phenomena was that the hills had been fissured by subterranean forces, and that the rivers had taken advantage of them to change their course. But close examination showed that these ravines were not fissures, but channels eroded in the rock, since the solid rock could often be traced unbroken across the very bed of the stream. And, after examining many ravines in different parts of the world, Mr. Beete-Jukes came to what then seemed the very startling conclusion that, except, perhaps, in districts recently convulsed by great earthquakes, there is no such thing as a glen, ravine, or valley occupying the upper portion of an open-mouthed fissure. On the contrary, in every case the whole space between the two sides of the valleys was once filled by rock, which has been gradually worn down and carried away. The very frequent presence of cascades and waterfalls in such ravines, formed by a continuous bed of hard rock crossing the stream, is itself sufficient to disprove the theory of fissures, in which case the whole bed would present a mass of fallen fragments, filled in with pebbles and sand; but this consideration seems never to have occurred to the upholders of the apparently obvious and easy theory of violent disruption.
It remains, however, to account for the very common phenomenon of rivers apparently going out of their way to cut a narrow passage through a hill, instead of following lower ground to a main valley or to the sea. Such in our own country are the small rivers Ouse and Cuckmere, which cut through the South Downs between
1 Student's Manual of Geology. By J. Beete-Jukes, 3rd ed. (edited by Archibald Geikie, F.R.S.), p. 450.
Brighton and Beachy Head, instead of following the low ground and reaching the sea between Eastbourne and St. Leonards; while the Avon, which flows through the gorge of St. Vincent's rocks at Clifton, might apparently have found a much easier way to the sea by a more northerly or a more southerly course. Mr. Jukes explains all these cases on the principle that the courses of almost all the rivers of a country were determined by the contour of the land when it first rose above the sea, the surface water seeking always the easiest course along the hollows and gentle slopes, without any regard to the nature of the rocks beneath. When once these streams had formed definite channels, it was almost impossible to alter them (except when diverted by lava streams or glaciers), because movements of elevation are so slow that the rivers can cut their way down as fast as the land rises up. Thus, the American geologists have proved that the Uintah Mountains were upheaved across the valley of the Green River after the course of that river was established, and that, as fast as they rose, the river cut through them, and now flows in a tremendous gorge or cañon. Another illustration of the permanence of river channels is afforded by the Moselle, which, although it flows at the bottom of a deep, narrow valley sunk in a nearly level plateau, winds about in great curves and deep horseshoe bends exactly like a stream flowing over a flat alluvial plain. No explanation of this can be given except that the river began its existence on a nearly level surface, and after it had established its course in the characteristic winding fashion of such streams, it has, in the course of long ages, cut its way deep down through the rock, and thus formed its present valley.
Now, every considerable area of continental land is made up of rocks and deposits of very unequal hardness and resisting power, from clays and sands to the various kinds of crystalline rock. Some of these can be dissolved and carried away by running water much more quickly than others; while rain, frost, and wind, also act upon their exposed edges very unequally. Hence arise the peculiar forms assumed by hills of different composition,
VOL. I. C
and hence the reason why valleys are in some parts very narrow and precipitous, in others wide and open. It is an invariable rule that hills and mountains are composed of the harder or less soluble rocks, the adjacent lowlands and valleys of the softer and more soluble. Hence, we see all great mountain ranges mainly composed of the older, hard, or crystalline rocks, while the lowlands, plains, and valleys are occupied by the newer and softer formations. In our own country the tertiary or secondary clays and sands are found in the lowland districts, while the more ancient and much harder rocks form the hills of Devonshire, Wales, the Lake District, and Scotland.
Keeping in mind the extreme inequality of the rate of denudation of different rocks, we are able easily to explain the apparently erratic course of so many rivers. When the streams originated they took their course along lines of least resistance, depending on the form of the surface, not on the nature of the rocks beneath the surface. Sometimes this course passed over ridges or bosses of very hard rock, buried perhaps hundreds, perhaps thousands, of feet deep. But the channels once fixed could not be altered, and when the bed of the stream reached this rock it cut down into it. Then, owing to the hardness of the rock, the river channel would be a gorge or ravine, while all around the softer rocks would be denuded by frost and rain, so that extensive areas would be lowered as fast as the stream cut its narrow channel through the hard rock, and was able to carry away the denuded material; while the gravel or sand thus carried down assisted in wearing away and cutting out the ravine through which it all had to pass. Hence, in the course of ages we should have the stream flowing over a wide lowland, perhaps on one side open to the sea, and then cutting straight across a mountain ridge, or even across an isolated hill entirely surrounded by lowlands.
Not very much time, geologically speaking, is required for such operations. Sir Charles Lyell describes a channel, cut by the river Simeto across a lava stream from Etna, which is over fifty feet wide and in some parts forty to fifty feet deep. The lava is not porous, but is a homo-
geneous mass of hard blue rock. Yet the date of the eruption which produced this lava stream is known to be 1603.1 But the most wonderful example of the power of water to denude and erode the hardest rocks is afforded by the great cañon of the Colorado river. This has been cut for about 400 miles to a depth of from 4,000 to 7,000 feet, mainly through masses of hard palæozoic rocks down to the archæan, and the whole of this vast operation has been performed in the latter half of the tertiary period. The formation of the river began, it is true, in very early tertiary times, but at that epoch the present surface was buried about 9,000 feet deep in secondary rocks, which have all been since denuded away, so that Captain Dutton estimates that the river has cut its channel on the whole through from 10,000 to 16,000 feet of mesozoic, carboniferous, and other ancient rocks, all during the tertiary period.2
Formation of the Inaccessible Valleys.
Keeping in mind these remarkable instances of denudation, let us turn to consider the probable origin of the remarkable valleys which have seemed to eminent geologists so peculiar as to need some special mode of origin; and we will take first the great rock-walled valleys of New South Wales, as being the most simple in their main features.
These are all excavated in sandstones and shales of the carboniferous system, though perhaps of mesozoic age. The strata are nearly horizontal, and, what is especially important, they are of very unequal degrees of hardness. The upper beds are usually conglomerates, and are so comparatively indestructible that isolated summits often imitate ruined castles. In places these beds are so hard that boring-tools will not penetrate them, while in other parts the rock is so incoherent that large blocks will break in pieces by falling over an embankment.3 We have here
1 Principles of Geology, vol. i., p. 353.
2 The Tertiary History of the Grand Cañon District, U.S. Geological Survey, 1882.
3 Remarks on the Sedimentary Formations of New South Wales. By the Rev. W. B. Clarke, F.R.S., F.G.S. Fourth ed, 1878, p. 72.
C 2
the essential conditions for the formation of vertical escarpments, since by the weathering away of the softer beds the harder strata above them remain unsupported and break off, and thus the vertical or sometimes overhanging character of the precipices is kept up.
If we look at a large-scale map of this part of Australia, we see that the rivers Grose, Cox, and other tributaries of the Nepean which drain the sandstone plateau, have great numbers of diverging branches which almost interlace with each other, as so often occurs among the streams of a nearly level well-watered district. Now, bearing in mind what has been said of the permanence of watercourses once formed, we can see that these many-branching streams must have flowed on the surface of the plateau at the epoch of its first elevation; that surface itself being perhaps considerably above the present surface, which has certainly been lowered by denudation during its long existence as dry land, probably during the whole of the tertiary period. From the time that these streams began to penetrate the sandstone plateau as far as the first hard bed, miniature cliffs would be formed by the wasting away of the softer beds beneath it, and the continual movement backward thus produced would widen the valleys till those of many of the smaller tributaries became united together. Thus age after age the valley would widen and deepen, always preserving its precipitous rock-walls due to the alternation of hard and soft layers.
The deepening of these great valleys would probably be aided by subterranean denudation due to the presence of salt and alum, which Mr. Clarke states are found at several places in these strata. The solution of these salts by percolating water would form cavities and water channels, and the subterranean streams would eat away the softer beds, forming caverns, the roofs of which would in time fall in, and the débris be gradually disintegrated by atmospheric agencies and then carried away by floods. This mode of denudation was seen actually at work by the late Sir George Grey, during his exploration of the Glenelg River in North-West Australia. He describes a nearly level table-land covered with numbers of sandstone pillars of
various grotesque shapes and some of them forty feet high. Hearing the sound of running water at a fissure among some of the rocks, he descended, and found a cavern supported by pillars of the same character as those above, with a small stream, which in the rainy season would become a torrent. Here, then, are ample causes to explain the formation of these great rock-walled branching valleys in the sandstone plateau; the remaining feature—that the rivers all escape through deep gorges often so narrow or so blocked up with rock-fragments as to be impassable —evidently depends on the fact that the outer escarpment of the plateau is formed of a series of harder rocks, and thus does not wear away laterally. In this respect they resemble those numerous gorges in the Alps which form the only outlet for high valleys of considerable extent, such as those of the Trient, the Reuss, and many others.
The difficulty as to whither the denuded material has gone, does not seem a great one, when we remember the many millions of years the process of denudation has been going on, with alternating epochs of greater rainfall producing more rapid-flowing streams and greater floods, by which the bulk of the sandy material would be carried out to sea, while the finer suspended matter would be deposited during wide-spreading floods on the valley bottoms and alluvial plains. The absence of great quantities of rock in the valleys themselves merely indicates that the degradation of the cliffs is now so comparatively slow that the fallen masses are worn down by atmospheric agency at about the same rate as they are reproduced.
Let us now see how the same general principles and the same denuding agencies will apply under the very different conditions which have prevailed in the district of the Yosemite. These differences are, mainly, the much loftier mountains and the very much greater extremes of climate; the recent occurrence both of glacial and of volcanic action on a large scale; and, lastly, the whole valley being excavated in granite instead of in sandstone rock.
The granite of the central and highest parts of the Sierra Nevada is flanked near the Yosemite with Silurian slates, lower by some triassic or Jurassic beds followed by enormous deposits of late tertiary gravels, which have been largely preserved from denudation by extensive flows of lava, the remnants of which form the numerous table-mountains so characteristic of the lower slopes of the Sierra. As granite is a metamorphic or igneous rock it can only have been formed deep down in the crust of the earth, where it was exposed to the action of great subterranean heat. It is therefore certain that, when first elevated to form the mass of the Sierra Nevada, it was everywhere deeply buried under Silurian and other palæozoic rocks, and not improbably under a further deposit of mesozoic age. These various beds, of an unknown thickness, must all have been denuded away before the granitic core was exposed, and during that process the main lines of the valleys must have been fixed, and the streams might have begun to cut their way into the granite substratum.
Although granite appears to be, and sometimes is, a very durable rock, it varies greatly in its power of resisting denudation, owing perhaps, in part, to the nature and thickness of the overlying rocks, beneath or among which it was forced up, and which in some cases determined the characteristic forms it assumes when exposed to atmospheric agencies. These forms are either rude cubical masses, as seen on some of our Dartmoor tors; peaks and pinnacles, as in some of the Alps of Dauphiné and in the Cathedral spires of the Yosemite; but, more commonly, rounded forms culminating in cones or almost perfect domes or hemispheres, as in the great domes of the Yosemite. (Fig. 7.) It is an interesting fact that all these forms occur also in the granite region of the Upper Kio Negro in Brazil The Cocoi Mountain forming the boundary between Brazil and Venezuela is a quadrangular or cubical mass of granite, about a thousand feet high, rising abruptly out of a great undulating plateau of the same rock. Others in the same region are conical or dome-shaped; and on the southern bank of the river Uaupes, about sixty miles from its mouth, is an isolated

FIG. 7.—THE NORTH DOME, YOSEMITE VALLEY.
dome-shaped mountain about a thousand feet high, of so regular an outline as to look like a gigantic half-globe. Now, it is evident that these cubical, hemispherical, and conical hills, rising out of a nearly level plateau which extends for several hundred miles around them in every direction, must owe their present position to the slow degradation by atmospheric agency of the vast masses of rock in which they were once buried, but whose destruction they have survived owing to their superior hardness or tenacity. It is true the rocks in Brazil have been subject to tropical rain and heat and to the powerful aid of tropical vegetation; but, on the other hand, the rocks of the Yosemite have been exposed to the even more powerful agencies of alternations of intense frost and great sun-heat, as well as of torrents formed by melting snows, and probably of occasional débâcles caused by bursting glacier lakes.
It is well known that granite often weathers very rapidly, sometimes becoming completely decomposed to a depth of twenty or thirty feet, so that it can be dug out with pick and spade. This process of decomposition is greatly facilitated by the action of carbonic acid either in air or water. Now, during the latter part of the tertiary epoch, there was a long period of volcanic action in the Sierra Nevada; and as both carbonic acid and many other powerful gases are emitted during eruptions, and also permeate the earth and are absorbed by the water, we should have all the conditions for the decomposition and denudation of the granite rocks. The alternations of temperature on the higher parts of the Sierra Nevada are very great. During the long bright Californian summer the action of direct sun-heat on the exposed rocks must be considerable, the air temperature in the Yosemite valley being usually over 80°, while at a height of 8,700 feet ice an inch thick is formed at night in June and July. In winter at such elevations—that of the present summit of some of the domes—the temperature must fall below zero of Fahrenheit every night. The alternate expansion and contraction produced by such changes of temperature are among the most powerful agencies in the splitting up
and decomposition of rocks. Small cracks thus produced receive water which freezes at night, and the crack is widened by the irresistible force of the ice wedge. It is by this agency that the final touches have been given to the Yosemite scenery, after all the softer and more decomposable portions of the rock had been removed by the ordinary modes of weathering. The huge domes and spires, and the subquadrangular mass of El Capitan, must be looked upon as intensely hard and compact cores of rock that remain after all the more friable masses that inclosed them have been removed. They show us the natural forms into which granite weathers, due perhaps to the mode in which it has originally cooled from the molten or plastic state. In the case of the dome the mass consists of concentric layers, probably of different densities, which peel off successively like the coats of a gigantic onion. On some of the domes we can see one of these coats partially removed, and the same thing was observed by myself in the dome-shaped mountains as well as in the smaller subglobular masses of granite in the Rio Negro.
The fact that the process of denudation, continued perhaps throughout the greater part of the tertiary period, has now eaten away all the more friable and soluble portions of the rocks which once occupied the site of the valley, leaving only those compact central masses which are hardly affected by ordinary atmospheric action, will account for what seemed such a great difficulty to Professor Whitney—the small amount of rock débris under the great precipices or in the valley generally. For the last few thousand years, probably, the amount of rock-falls has been comparatively small, so that it barely equals the rate at which atmospheric agencies, aided by vegetation, break up and decompose the fallen masses, which then, in the form of the coarse granitic sand that constitutes the surface soil in all the drier portions of the valley, is gradually carried by wind, rain, and melting snow into the river, and ultimately into the great bay of San Francisco. That some considerable amount of decay is still going on in these giant cliffs is evident, not only
from the rock-falls that actually occur every year, but from the numerous places where great flakes or jutting blocks can be observed in every stage of detachment from the parent rock. These fallen masses, however large, are at once subject to fresh causes of decay. Almost all their surfaces are exposed to atmospheric action or to expansion and contraction by heat and cold. Every crack and cranny is seized upon by vegetation—first the lowly herb, then the shrub, later the tree, whose roots penetrate the minutest fissure, eat away the surface, or even split off portions by the power of growth. And though in the life of a man a block may seem unchanged, in a few thousand years it may have entirely disappeared; and such a lapse of time probably bears a less proportion to the period occupied by the valley's formation, than does a single hour to the life of a man.
It has now, I think, been shown that the two remarkable valleys here described do not owe their exceptional physical features to any catastrophic or unusual mode of origin. Every characteristic they possess is fully explained by that simple theory of earth sculpture by atmospheric agency which has been found applicable to the solution of similar problems in all other parts of the world. This theory does not, of course, imply that subterranean movements have no part in determining the direction or hastening the excavation of some valleys, but only that they have in no case produced the valleys themselves. Many examples can be pointed out in which valleys follow for a certain distance lines of fault, of the junction of different strata, or of the fractured summit of an anticlinal; but the explanation of these cases is, probably, that during elevation above the sea, wave-action produced slight hollows along these several lines of weakness, and that the hollows thus formed were occupied by the primitive rivulets as their line of least resistance when flowing towards the ocean. But these cases are very few as compared with those of valleys which pay no regard whatever to the geological features of the under-crust, but which cross over faults and outcrops, and break through transverse hills and mountain ranges, as if the
causes which determined their direction of flow were of an altogether different nature. And as regards what used to be considered the most striking cases of "valleys of disruption"—the narrow defiles and gorges like those of the Trient and the Reuss—it may now be affirmed, that in no single instance which has been carefully examined has any evidence of an open fissure been discovered, while in most cases there is the clearest proof that the gorges in question have been wholly excavated by the action of running water under conditions which are explained in Chapter V.
It was for the purpose of bringing clearly before non-geologic readers the total inaccuracy of the popular view —that every rock-walled valley or deep Alpine gorge has had its origin in some great "convulsion of nature"—and to impress upon such readers the grand but simple theory, which we owe mainly to the late Sir Charles Lyell, of the efficiency of causes now in action in producing the varied contours of the earth's surface, that this account of some of the most remarkable of known valleys has been written.
CHAPTER II
THE PERMANENCE OF THE GREAT OCEANIC BASINS
THAT the great Oceanic Basins, as well as the land areas of the globe, have persisted throughout a large portion, if not the whole, of known geological time, is a proposition which has been accepted by writers of such eminence, and is supported by so many distinct lines of evidence, that it seemed likely to become one of the established teachings of geology. Professor Dana was led to it by a study of the development of the North American Continent; Darwin upheld it from his study of Oceanic Islands, and the facts he adduced have since been strengthened by the discovery that the two supposed exceptions to the generalisation that no ancient sedimentary rocks occur on such islands—Rodriguez and St. Paul's Rocks—are no exceptions at all. Two successive heads of the Geological Survey of Great Britain, Sir Andrew Ramsay and Sir Archibald Geikie, have advocated similar views; while Mr. John Murray, of the Challenger, holds that the vast mass of evidence now accumulated as to the nature of the deposits on the floors of the great oceans, indicates that they are distinct in character and origin from any of the widespread formations which make up the series of the sedimentary rocks. Coming to the subject from a totally different point of view, that of the physicist and mathematician, the Rev. Osmond Fisher arrives at similar results. In the latest issue of his important work, "Physics of the Earth's Crust," he gives as one of his conclusions—"and
lastly, that the great oceanic and continental areas have never changed places"; and, in the summary of the whole work, he says: "The occupation of an entire hemisphere by one great ocean is a remarkable circumstance, and we have seen reason for believing that this is a very ancient division of the surface, and that it is probably a mistake to suppose that every part of it has been sometimes raised above the sea, and sometimes depressed beneath it. The truth seems to be, that the region subject to these alterations of conditions does not extend very far away from the present coast lines."
When studying the causes which have brought about the geographical distribution of animals, I was compelled to deal with this question, because I found that it had been the custom of many writers to solve all anomalies of distribution by the creation of hypothetical lands, bridging across the great oceans in various directions and at many different epochs; and, having arrived at the conclusion that the distribution of organisms could be more harmoniously and consistently explained without such changes of sea and land, which usually created greater difficulties than those they were intended to explain, I gave, in my Island Life, a brief statement of the evidence which appeared to me to render such changes exceedingly improbable. This evidence was mainly a summary of the facts and arguments adduced by the eminent men referred to above, and to this I added in my Darwinism a difficulty founded on mechanical considerations which seemed to me to furnish a preliminary reason why we should not accept the doctrine of the interchange of continental and oceanic areas without very clear and cogent reasons. Since then some other arguments of this nature have occurred to me, and as the theory of permanence has been recently attacked, by Mr. W. T. Blanford in his presidential address to the Geological Society in 1890, and by Mr. Jukes-Browne in his "Building of the British Isles," it may be as well to consider these difficulties, which suggest, in my opinion, a very powerful argument against the interchange of oceanic and continental areas, and one which has the advantage of not requiring any
knowledge of the higher mathematics in order to estimate its validity.
And, first, it is necessary to clear away some misconceptions as to the proposition I really uphold, since arguments have been adduced which in no way affect that proposition. Thus, Mr. Jukes-Browne quotes Professor Prestwich as saying, "It is only the deeper portions of the great ocean troughs that can claim the high antiquity now advocated for them by many eminent American and English geologists." But this is all that is claimed. For practical purposes I at first took the 1,000-fathom line as, generally and roughly, indicating the separation between the oceanic and the continental areas, because at that time it did accurately divide the continental from the oceanic islands, as defined by a combination of geological and biological characters. It has, however, been since shown that two ancient continental islands—Madagascar and New Zealand—are separated from their respective continents by depths of more than 1,000 fathoms. We must, therefore, go as far as the 1,500-, or, perhaps, in a few cases, to the 2,000-fathom line, and this will surely mark out "the deeper portions of the great ocean basins," since only isolated areas exceed 3,000 fathoms.
Now, if we look at the deep ocean basins marked out by the 2,000-fathom line, not on Mercator's projection which greatly exaggerates the extent of the shallower portion situated in the temperate and polar regions, but on an equal-area projection, such as the map which illustrates Sir J. Murray's paper,1 we shall see that by far the larger part of all the great oceans are included by this line, and that, for the purpose of indicating the isolation of the continents from each other throughout the equatorial and most of the temperate zones, there is very little to choose between the 1,000-fathom or the 2,000-fathom boundary. The latter, however, allows more scope for possible land extensions between the three southern continents and the Antarctic lands, which, during mild epochs, and by means of intervening islands, may, perhaps, have served as a
1 "On the Height of the Land and the Depth of the Ocean," Scottish Geographical Magazine, 1888.
means of communication between these continents. All that is necessary to maintain, therefore, is that existing continents with their included seas, and their surrounding oceanic waters as far as the 1,500-fathom, or in some extreme cases the 2,000-fathom line, mark out the areas within which the continental lands of the globe have been built up; while the oceanic areas beyond the 2,000-fathom line, constituting, according to Mr. Murray's data, 71 per cent. of the whole ocean, have almost certainly been ocean throughout all known geological time.1
It will now be seen that this is a problem which deals with the very broadest contrasts of the earth's surface, and that its fundamental data are on so vast a scale as not to be materially affected by the smaller details of physical geography, or by differences of opinion as to the exact meaning of certain terms. Whether a particular island is more correctly classed as oceanic or continental, whether a certain portion of the ocean should be placed within the oceanic or the continental area, and whether certain rocks were formed in very deep or in comparatively shallow water, are of slight importance, except in so far as they may throw light on the real question, which is, whether the vast expanses of ocean beyond the 1,000-fathom line (as shown by the map in Chap. XII. of my Darwinism), forming about 92 per cent. of the whole oceanic area, have ever been occupied, or extensively bridged over, by continental land. It is towards the solution of this great problem that I now propose to submit certain general considerations which appear to me to lie at the root of the whole matter.
Comparison of Oceanic and Continental Masses.—In the paper already referred to, Sir John Murray has carefully estimated both the area of the land and of the water on the earth's surface, and their bulk as deduced from the best available data. Taking the whole area of the globe as 100, he finds the land surface to be 28, the water surface 72. But the mean height of the land above sea-level
1 Sir J. Murray gives his results for oceans and inland seas together. The above percentage refers to the oceans only as usually understood; but the difference is not great.
is 2,250 feet, while the mean depth of all the oceans and seas is 12,456 feet. In this estimate, however, all the inland seas and shallow coast waters are included, and as these, at least as far as the 100-fathom line, are universally admitted to be within the "continental area," we omit them in our estimate of the mean depth of the oceans proper, which are thus brought to something over 15,000 feet, or nearly seven times as much as the mean height of the land.
The accompanying diagram (taken from my book on Darwinism) will better enable the reader to appreciate these proportions, which are of vital importance in the problem under discussion. The lengths of the two parts of the diagram are in proportion to the areas of land and
Diagram of proportionate mean height of Land and depth of Oceans.

FIG. 8.
ocean respectively, the vertical dimensions, although for distinctness on a greatly exaggerated scale, showing the comparative mean height and depth with sufficient accuracy. It follows that the areas of the two shaded portions are proportional to the bulk of the continents and oceans respectively.
The mean depths of the several oceans and the mean heights of the several continents do not differ enough from each other to render this diagram a very inaccurate representation of the proportion between any of the continents and their adjacent oceans; and it will therefore serve, roughly, to keep before the mind what must have taken place if oceanic and continental areas had ever changed places. It will, I presume, be admitted that, on any large
scale, elevation and subsidence must nearly balance each other, and, thus, in order that any area of continental magnitude should rise from the ocean floor till it formed fairly elevated dry land, some corresponding area must sink to a like extent. But if such subsiding area formed a part or the whole of a continent, the land would entirely disappear beneath the waters of the ocean (except a few mountain peaks) long before the corresponding part of the ocean floor had approached the surface. In order, therefore, to make any such interchange possible, without the total disappearance of the greater portion of the subsiding continent before the new one had appeared to take its place, we must make some arbitrary assumptions. We must suppose either that when one portion of the ocean floor rose, some other part of that floor sank to greater depths till the new continent approached the surface, or, that the sinking of a whole continent was balanced by the rising of a comparatively small area of the ocean floor. Of course, either of these assumed changes are conceivable and, perhaps, possible; but it seems to me that they are exceedingly improbable, and that to assume that they have occurred again and again, as part of the regular course of the earth's history, leads us into enormous difficulties. Consider, for a moment, what would be implied by the building up of a continent the size of Africa from the mean depth of the ocean. By comparing the area of Africa with that of the whole of the land, and the depth of the ocean with the mean height of the land, we shall find that if all the land of the globe above sea-level could be transferred to mid-ocean, it would not be sufficient to form the new continent, but would still leave it nearly 2,000 feet beneath the surface.
It thus appears that, if the elevation of the ocean floor, and the corresponding sinking of whole continents, constitute a portion of the regular change and development of the earth's surface, there would be not only a chance but a very great probability of entire continents disappearing beneath the waters before even the smallest new continent had risen to take their place. Even the total disappearance of all the large land masses might easily
VOL. I. D
happen; for we see from the diagram that they might one after the other disappear with a corresponding rise of the adjacent portion of the ocean bed and still leave the ocean over the whole earth almost as deep as it is now. But, as will be shown further on, the geological record, imperfect as it is, teaches us that no such general submergences have ever taken place.
Contour of the Ocean Floor as indicating Permanence.—Before extensive soundings revealed the depth of the ocean and the form of its floor, it was supposed that it would exhibit irregularities corresponding to those of the land, such as mountain-ranges, great valleys, escarpments, ravines, etc. But we now know that the main characteristic of the ocean floor is, that it is a vast slightly undulating plain, the slopes rarely exceeding a hundred feet in a mile except near the margins of the continental areas, while usually the gradients are so slight that they would be hardly perceptible to the eye. Contrast this with the forms of all mountain ranges whose general rise for long distances is often several hundred feet in a mile, while slopes at angles of from 20° to 60° are by no means uncommon.
Now if we suppose that considerable portions of the ocean depths have been formed by the subsidence of continents, we should certainly expect to find some indication of those surface features which characterise all the continents, but which appear to be absent from all the deep oceans. In order to account for the actual contours of the ocean on this theory, we must suppose that, during subsidence, all the mountain ranges, peaks, valleys, and precipices were reduced to an almost uniform level surface by marine denudation, which, unless the process of subsidence were incredibly slow, seems most improbable. Mr. Jukes-Browne, however, does not hold the view that they have been thus denuded, for he approvingly quotes Mr. Crosby as saying that—"the oceanic islands are, of course, merely the tops of submerged mountains, and it is only with the highest points of continents that they can properly be compared." If this is correct, then we ought to find in the vicinity of such islands all the chief features
of submerged mountain ranges—precipices, deep valleys and ravines, arranged in diverging groups as they always occur in nature. But in no single case, that I am aware of, have any such features been discovered.
But a still greater difficulty remains to be considered. If oceanic and continental areas are interchangeable, it can only be because the very same causes (whatever they may be) that produce elevations and subsidences in the one produce them also in the other, and at first sight it appears probable that this would be the case. But if these causes have been at work upon the ocean floor throughout all geological epochs, they would have produced irregularities of surface not less but far greater than on subaerial land. This must be so, because subaerial denudation continually neutralises much of the effect of upheaval in the continental areas, while in the ocean depths no such cause or anything analogous to it is in operation.
The forces which have been at work in every mountainous region have sometimes tilted up great masses of rock at high angles, upheaved them into huge anticlinal curves, or crushed them by lateral pressure into repeated folds, which in some cases appear to have fallen over so as to reverse the succession of the strata. But, notwithstanding these various forms of upheaval, involving vertical displacements which are sometimes several miles in extent, the surface of the land usually shows no corresponding irregularities, owing to the fact that subaerial denudation has either kept pace with upheaval or has even exceeded it, so that the position of an anticlinal ridge may be, and often is, represented by a valley. Now, if we suppose that similar upheaving forces have been at work on that portion of the earth's surface forming the bed of the ocean where there are no such counteracting agencies, we should expect to find irregularities in the ocean floor far greater than those which occur upon the land surface.
Still more difficult to explain would be the absence of precipitous escarpments due to faults, which, though frequently showing an upthrow or downthrow of the strata to the amount of many hundreds and sometimes many
D 2
thousands of feet, rarely exhibit any difference of level on the land surface, owing to the fact that subaerial denudation has kept pace with slow and intermittent elevation. But in the ocean depths no such denudation is going on; and we can therefore only account for its very uniform surface on the supposition that it is not subject to the varied and complex subterranean movements which have certainly acted within the continental areas throughout all known geological time.1
Similar Range of the Geological Record in all the Continents.—There is one other general consideration which indicates the permanence and continuity of the Continental Areas, and which renders it very difficult, if not impossible, to suppose that they have ever changed places with the great oceans. It is, that on all the present continents we find either the same or a closely parallel series of geological formations, from the most ancient to the most recent. Not only do we find Palæozoic, Mesozoic, and Cainozoic rocks everywhere present, but, in proportion as the continents are explored geologically, we find a tolerably complete series of the chief formations. From Laurentian to Carboniferous and Permian, from Trias to Cretaceous, and from Eocene to Quaternary, the geological series appears to be fairly represented, not in continents only but also to a considerable extent in the large continental islands such as Great Britain and New Zealand.
Now this is certainly not what we should expect if the present continental areas had, at different epochs, risen out of the deep oceans. In that case some would have commenced their geological history at a later period than others, having either a late Palæozoic or some Mesozoic formation, or even an early Tertiary for their very lowest stratified rock. Others, which had become oceanic for the first time at a later epoch, would exhibit an enormous gap in the series, either several of the Mesozoic formations, for example, being absent, or some considerable portion of
1 The Rev. O. Fisher has arrived at the same conclusion from his own researches. He says: "The compression which has caused the thickening accompanied by corrugation, such as characterises most elevated tracts, is properly a continental phenomenon, and has no analogue beneath the oceans." L.c., p. 253.
both Palæozoic and Mesozoic, or Mesozoic and Tertiary. This would necessarily be the case, because we cannot believe that so vast a change as the subsidence of an entire continent till its site became a deep ocean, and its subsequent elevation till it again became dry land, could possibly be effected in any less extended periods, if at all.
Whenever such gaps, or smaller ones, now occur locally, they are generally held to imply the existence of terrestrial conditions, as in the case of China, which, according to Richthoven, has been continental since the Carboniferous epoch. In many cases there is distinct evidence of such conditions in lacustrine or freshwater deposits, dirt-beds, &c. But if a gap of such vast extent, both in space and time, as that here referred to were caused by the interchange of a continent and a deep ocean, the fact that it was so produced would be clearly evidenced by an almost uniform deposit either of organic or clayey ooze, similar to those now everywhere forming over the oceanic area. Even if we make the fullest allowance for denudation during elevation, sufficient indications of so widespread a formation should be detected. Such a deposit would, in fact, have every chance of being largely preserved, because, long before it rose to the level where it would be subject to denudation by waves or currents, it would, almost everywhere, be overlain by a series of shore deposits, and where-ever these latter were preserved on the land surface the oceanic formation would necessarily be found under them. That no such enormous deficiency in the geological series characterises any of the continents, and that no widespread deposit of organic or clayey ooze at some definite horizon has been anywhere detected, though such a deposit must have been formed and largely preserved if the whole or any considerable part of a continent had risen from ocean depths at any period of its history, constitute of themselves very strong arguments against any such interchange of oceanic and continental areas having occurred.
Summary of the Argument.
I have now shown that there are three distinct groups of phenomena which are either altogether inconsistent
with any general interchange of oceanic and continental areas, or which render it exceedingly difficult to understand how such interchange could have been brought about. These phenomena are:—(1) The enormous disproportion between the mean height of the land and the mean depth of the ocean, which would render it very difficult for new land to reach the surface till long after the total submergence of the sinking continent. (2) The wonderful uniformity of level over by far the greater part of the ocean floor, which indicates that it is not subject to the same disturbing agencies which throughout all geological time have been creating irregularities in the land-surface, irregularities which would be far greater than they are were they not continually counteracted by the lowering and equalising effects of subaerial denudation. (3) The remarkable parallelism and completeness of the series of geological formations in all the best known continents and larger continental islands, indicating that none of them have first risen from the ocean floor during any portion of known geological history, a conclusion enforced by the absence from any of them of that general deposit of oceanic ooze at some definite horizon, which would be at once the result and the proof of any such tremendous episode in their past history.
I submit that these facts, and the conclusions to be logically deduced from them, form a very powerful, if not a conclusive argument as against those who maintain the interchange of continents and oceans as a means of explaining certain isolated geological or biological phenomena; such, for instance, as the much-disputed origin of the chalk, or the supposed necessity for land-communication to explain the distribution of certain groups of mammals or birds, reptiles or fishes in remote geological times. Before postulating such vast revolutions of the terrestrial surface in order to cut the gordian knot of difficulties which may be mainly due to imperfect knowledge, it will be necessary to show that the considerations here adduced, as well as the great body of facts which have caused many eminent geologists, naturalists, and physicists to hold the doctrine of oceanic permanence, are either illogical or
founded on incorrect data. For, surely, no one will suggest that so vast a problem of terrestrial physics can be held to be solved till we have exhausted all the evidence at our command, and have shown that it largely preponderates on one side or the other.
The present article is intended to supply a hitherto unnoticed class of arguments for oceanic permanence, and these must of course be taken in connection with the other evidence which has been summarised in the sixth chapter of the writer's Island Life, and also with the admirable "Summary" of the purely physical argument in the second edition of Mr. O. Fisher's Physics of the Earth's Crust. It is certainly a remarkable fact that writers approaching the subject from so many distinct points of view—as have Professor Dana, Mr. Darwin, Sir Archibald Geikie, Dr. John Murray, Rev. O. Fisher, and myself—should yet arrive at what is substantially an identical conclusion; and this must certainly be held to afford a strong presumption that that conclusion is a correct one.
CHAPTER III
OUR MOLTEN GLOBE
FEW scientific inquiries excite greater interest than those recent researches which have so greatly extended our knowledge of the stars and nebulæ, whether by determining the direction and velocity of their motions, or ascertaining their physical constitution and probable temperature. In comparison with this considerable amount of knowledge of such distant bodies, it seems strange that so little comparatively is known of the structure and internal constitution of the globe on which we live, and that much difference of opinion should still exist on the fundamental question whether its interior is liquid or solid, whether it is intensely hot or comparatively cool. Yet the definite solution of this problem is a matter of the greatest theoretical interest, since it would not only elucidate many geological phenomena, but might possibly serve as a guide in our interpretation of appearances presented by the other planets and even by more remote bodies; while it is not unlikely that it may soon become a practical question of the highest importance, inasmuch as it may lead us to the acquisition of a new source of heat, in many ways superior to that produced by the combustion of fuel, and practically inexhaustible.
It is only during the present century that facts have been accumulating in various directions, bearing more or less directly on the question of the earth's internal condition. These have been partially dealt with, both by geologists and by physicists; but the problem is such a
complex one, and the evidence of so varied a nature and often so difficult to interpret, that the conclusions reached have been usually doubtful and often conflicting. This seems to have been due, in part, to the fact that no properly qualified person had, till quite recently, devoted himself to a thorough study of the whole subject, taking full account of all the materials available for arriving at a definite conclusion, as well as of the various groups of phenomena which such a conclusion must harmonize and explain. But for many years past a good practical geologist, who is also an advanced mathematician—the Rev. Osmond Fisher—has made this subject his speciality, and in a most interesting volume, of which a second and carefully revised edition, with an appendix, has been recently published, he has brought together all the facts bearing on the problem, and has arrived at certain definite conclusions of the greatest interest. The object of this chapter is to give a popular account of so much of his work as bears upon the question of the thickness and density of the earth's crust and the constitution of the interior.1
The Argument form Internal Heat.
We will first consider the nature of the evidence in favour of the view that, below a superficial crust, there is a molten or highly heated substratum. The existence of volcanoes, geysers, and hot springs irregularly scattered over the whole surface of the globe, and continually ejecting molten rock, ashes, mud, steam or hot water, is an obvious indication of some very widespread source of heat within the earth, but of the nature or origin of that heat they give little positive information. The heat thus indicated has been supposed to be due to many causes, such as the pressure and friction caused by contraction of the cooling crust, chemical action at great depths beneath the surface, isolated lakes of molten rock due to these or to unknown causes, or to a molten interior, or at least a
1 Physics of the Earth's Crust, by the Rev. Osmond Fisher, M.A., F.G.S. Second edition, altered and enlarged. Macmillan and Co., 1889. With an Appendix, 1891.
general substratum of molten matter between the crust and a possibly solid interior. The first two causes are now generally admitted to be inadequate, and our choice is practically limited to one of the latter.
There are also very important evidences of internal heat derived from the universal phenomenon of a fairly uniform increase of temperature in all deep wells, mines, borings, or tunnels. This increase has been usually reckoned as 1° F. for each 60 feet of descent, but a recent very careful estimate, by Professor Prestwich, derived from the whole of the available data, gives 1° F. for every 47.5 feet of descent. It is a curious indication of the universality of this increase that, even in the coldest parts of Siberia, where the soil is frozen to a depth of 620 feet, there is a steady increase in the temperature of this frozen soil from the surface downwards. Much has been made by some writers of the local differences of the rate of increase, varying from 1° in 28 feet to 1° in 95; and also of the fact that in some places the rate of increase diminishes as the depth becomes greater. But when we consider that springs often bring up heated water to the surface in countries far removed from any seat of volcanic action, and the extent to which water permeates the rocks at all depths reached by man, such divergences are exactly what we might expect. Now this average rate of increase, if continued downwards, would imply a temperature capable of melting rock at about twenty miles deep, or less, and we shall see presently that there are other considerations which lead to the conclusion that this is not far from the average thickness of the solid crust.
Before going further it will be well to consider certain objections to this conclusion, which for a long time were considered insuperable, but which have now been shown to be either altogether erroneous or quite inconclusive. In Sir Charles Lyell's Principles of Geology, Mr. Hopkins is quoted as having shown that the phenomenon of the precession of the equinoxes, due to the attraction of the sun and moon on the equatorial protuberance, requires the
1 In a recent deep boring at Wheeling, Virginia, the rate of increase was found to be greater as the depth increased.
interior of the earth to be solid, or at least to have a crust not much less than one thousand miles thick. This view was supported by Sir William Thomson and other eminent mathematicians, and so great was the faith of geologists in these calculations that for nearly forty years the theory of the earth's internal liquidity was almost wholly abandoned. But this argument has now been shown to be erroneous by the more complete investigations of Professor George Darwin, while Sir William Thomson (now Lord Kelvin) has recently shown experimentally that a rotating liquid spheroid behaves under stresses as if it were a solid. Another difficulty arises from the phenomena of the tides. It has been argued that, if the interior of the earth is liquid tides will be formed in it which will deform the crust itself, and thus, by lifting the water up with the land, do away with any sensible tides in the ocean. But Mr. Fisher has pointed out that this conclusion rests on the assumption that the liquid interior, if it exists, is not an expansible fluid; and he shows that if this assumption is incorrect it is quite possible that little or no deformation would be caused in the crust by tides produced in the liquid interior; and he further maintains, as we shall see presently, that all the evidence goes to prove that it is expansible. Moreover, in a late paper, he claims to have proved that even the deformation of the crust itself would not obliterate the ocean tides, but would diminish them only to the extent of about one-fifth.1
There remain the geological objections founded on the behaviour of volcanoes, which is supposed to be inconsistent with a liquid interior as their effective cause. We have, for instance, the phenomenon of a lofty volcano like Etna pouring out lava from near its summit, while the much lower volcanoes of Vesuvius and Stromboli show no corresponding increase of activity; and the still more extraordinary case of Kilauea, on the lower slopes of Mauna Loa, in the Sandwich Islands, at a height of about 3,800 feet, whose lake of perennial liquid lava suffers no alteration of level or any increased activity when the parent mountain is pouring forth lava from a height of 14,000
1 Proceedings of the Cambridge Philosophical Society, 1892.
feet. Again, it is argued that if the igneous products of volcanoes are derived from one central reservoir there ought to be a great similarity between them, especially between those of the same district. But this is not the case, an example being the Miocene lavas of Hungary and Bohemia, which are of a totally different character, chemically, from each other. But although the molten interior of the globe may be the common source of the heat which causes volcanic eruptions, it by no means follows that the whole, or any large portion, of the matters ejected from volcanoes are derived from it; and it is a remarkable indication of the probable truth of Mr. Fisher's theory, that, as will be shown further on, it entirely removes the two geological difficulties here noticed. At the same time it explains other geological phenomena of a striking character which the theory of solidity altogether fails to account for, as will be now briefly indicated.
The Argument from Subsidence under Deposition of Sediment.
It has long been known to geologists that the series of sedimentary rocks, ancient as well as modern, afford repeated examples of great piles of strata hundreds, or even thousands of feet thick, which throughout present indications of having been formed in shallow water, and which therefore imply that as fast as one bed was deposited it sank down, and was ready to receive another bed on the top of it. As an example we may refer to the Palæozoic rocks of the Alleghany Mountains, which are not less than 42,000 feet thick; yet the lowest of these strata, the Potsdam sandstone, was not deposited in a deep sea, but evidently in shallow water near shore, several of the beds exhibiting distinct ripple markings, and the same is the case with the highest strata found there—the carboniferous. On this point Sir Archibald Geikie remarks:—
"Among the thickest masses of sedimentary rocks—those of the ancient Palæozoic systems—no features recur more continually than the alternation of different sediments, and the recurrence of surfaces covered with well-preserved ripple-marks, trails and burrows of
annelides, polygonal and irregular desiccation marks like the cracks at the bottom of a sun-dried muddy pool. These phenomena unequivocally point to shallow and even littoral waters. They occur from bottom to top of formations which reach a thickness of several thousand feet. They can be interpreted only in one way, namely, that the formations in question began to be laid down in shallow water; that during their formation the area of deposit gradually subsided for thousands of feet, yet that the rate of accumulation of sediment kept pace on the whole with this depression; and hence that the original shallow-water character of the deposits remained after the original sea-bottom had been buried under a vast mass of sedimentary matter."
Coming now to the other end of the geological record, we find in the deltas of existing rivers an exactly similar phenomenon. At Venice a boring of 400 feet deep was entirely in modern fluviatile mud, the bottom of which was not reached; and at four separate depths, one of them near the bottom, beds of turf or of vegetable matter were passed through, showing, as Sir Charles Lyell observes, "that a considerable area of what was once land has sunk down 400 feet in the course of ages."1 At Zagazig, on the eastern border of the Nile delta, borings have been made for the Royal Society, and have not found rock at a depth of 345 feet. In the delta of the Mississippi a well at New Orleans, 630 feet deep, passed entirely through sands and clays, with fresh-water shells of living species. Again, in the delta of the Ganges, at Calcutta, a boring 481 feet deep was entirely through beds of sand, peat, gravel, and other alluvial or fresh-water deposits. This remarkable concurrence of testimony from so many parts of the world and from different geological periods, indicates a general law of subsidence so uniformly coinciding with deposition, and so regularly keeping pace with it, that we can hardly avoid the conclusion that the two phenomena are connected; and the most reasonable explanation seems to be that the deposit of matter in a shallow sea directly causes the depression of that bottom by its weight. Such depression is quite intelligible on the theory of a thin crust resting or floating on a liquid substratum, but is quite unintelligible on the supposition of a solid globe, or of a crust several
1 Principles of Geology, 11th ed., vol. i., p. 422.
hundred miles thick. It is only reasonable to suppose that depression thus caused must be accompanied by a corresponding elevation of some other area, and as there must always be an adjacent area from which an equivalent weight of rock has been removed by denudation, we should expect the elevation to occur there; and many geologists believe that there is direct evidence of elevation wherever areas are being rapidly denuded.
In a very interesting letter to Nature (Dec. 5th, 1889) Mr. J. Starkie Gardner states that he has actually observed the results of denudation to be of this character. He says:—
"The immediate effect of cutting down cliffs, say of 100 feet in height, and removing them by wave-action, is to relieve the pressure at their base; and I claim that, wherever I have excavated for the purpose of collecting under such conditions, I have found a decided slope inwards away from the sea, if the strata were at all horizontal, no matter what direction their general slope might be at a distance from the sea margin. But on the beach, a little way from the base of the cliffs, the slope is, on the contrary, towards the sea.… This appears to me to be simply because the relief from pressure has made the beach-line the crown of a slight arch, and an arch that continues to grow and travel."
Hence he concludes that—
"Whether we look at the past or the present, we seem to see evidence of a crust resting in equilibrium on a liquid layer, and sensitive to even apparently insignificant readjustments of its weight."
The physical and geological phenomena of which an outline sketch has now been given, all point unmistakably to a thin crust of various rocks resting on a molten substratum; but there are certain difficulties and objections which require a fuller discussion. In order to remove these difficulties and answer these objections, we must, with the aid of Mr. Fisher's work, go more deeply into the question, and we shall then find that, by means of some of the most refined inquiries of modern physicists, we are able to obtain so much additional information as to the peculiarities of the crust and of the substratum, that most, if not all, of the alleged difficulties will be found to disappear.
The Argument from Variation of the Force of Gravity.
It is well known that mountains attract the plumb-line, and thus render latitudes determined by its means, or by a spirit or mercurial level, inaccurate in their vicinity. During the trigonometrical survey of India the amount of this error was carefully determined in several localities near mountains, but a discrepancy appeared. When the mass of the Himalayas was estimated and its attraction calculated, it was found to be more than the observed attraction. The same thing had occurred in the original experiment by Maskelyne at Schehallion in Scotland; and a similar deficiency in the error produced was noticed by Petit in the case of the Pyrenees. Many attempts were made to explain the discrepancy, but that which was advanced by the late Sir G. B. Airy seems best to account for all the phenomena, and is that adopted by Mr. Fisher. It is, that every mountain mass on a continent has a much larger mass projecting beneath the crust into the liquid substratum, exactly as an iceberg has a larger mass under the water than above it. Sir G. B. Airy argued that, whether the crust were ten miles or a hundred miles thick, it could not bear the weight of such a mass as the Himalayan and Tibetan plateaus without breaking from bottom to top, and receiving support by partially sinking into the liquid mass. The best experiments show that the proportionate densities of most rocks in a solid and a liquid state are approximately as ice is to water, and thus no mountain masses can be formed, whether by lateral pressure or other agency, without a corresponding protuberance forming below to keep the crust in equilibrium. It is this displacement of the denser substratum by the less dense "roots of the mountains" that leads to the total attraction of such mountains being less than they otherwise would be. In our author's words—"the roots of the mountains can be felt by means of the plumb-line."
Still more important and interesting are the revelations afforded by the pendulum, since they not only support the interpretation of the plumb-line experiments above given,
but furnish additional material for estimating the varying thicknesses and densities of the earth's crust. The rate of vibration of a pendulum of constant length depends upon the force of gravity at the place, and thus variations in that force can be determined with considerable accuracy. Taking the number of vibrations in a day of a seconds pendulum at the equator and at the sea-level as 86,400, the number of vibrations at any other latitude can be calculated on the theory that the earth is a perfect spheroid of revolution; and geodetic observations show that it has such a form. At any elevated station, whether on an isolated mountain or on an extensive plateau, the pendulum will vibrate more slowly on account of its greater distance from the centre of gravity of the earth, while it would vibrate more quickly on account of the additional attraction of the elevated mass immediately beneath and around it. These effects can be calculated, and the balance of the two, applied to the normal rate for the latitude, will give the theoretical rate due to the position and altitude of the station. Experiments were made at more than twenty stations in India, varying from the sea-level to over 15,000 feet above it, and at all the higher stations there was a deficiency of the observed as compared with the calculated number of vibrations, of from one to twenty-four vibrations in the twenty-four hours. In such delicate observations there were of course some irregularities, but the fact of a greater deficiency at the higher levels came out very clearly, and could be explained only by a deficiency of subterranean density due to the roots of the mountains displacing a denser substratum, as in the case of the plumb-line experiments.
Before leaving this subject of the "roots of mountains," it will be well to refer to a remarkable corroboration of their actual existence by evidence of a quite different kind. It has already been pointed out that the rate of increase of underground temperature would, if continued downwards till the heat equalled the melting point of rock, give a mean thickness of the crust of about twenty miles. But in places where the crust is so much thicker, as it is supposed to be under mountains, the rate of increase
should be much less, because the lower level of the crust in contact with the liquid substratum must always be at about the same temperature—that of melting rock. This is found to be the case; the rate of increase at the St. Gothard tunnel, where the observations were most complete, being 1° F. in eighty-eight feet, and the corresponding thickness of the crust thirty-seven miles. This is certainly a remarkable confirmation of the other observations, and of the theory of mountains being supported in approximate equilibrium by means of vast protuberances into the liquid substratum beneath.
The general result of the whole series of experiments with the pendulum shows that gravity is normal at the sea-level both over land and sea, and thus proves that the surface of the globe is in a state of equilibrium. The measures of the force of gravity over the oceans have been necessarily taken on islands, and have led to a curious discovery. The pendulum experiments on oceanic islands such as the Galapagos, Ascension, St. Helena, Bourbon, Guam, and others, all show an increase in the force of gravity, which, on the average, is very nearly accounted for by the subaqueous mass of land displacing water of less than half the density of rock. Hence it is concluded that these islands or island-mountains do not have "roots" as do those on continents; and the same thing occurs with isolated volcanoes on continents, the attraction of Fujisan in Japan being exactly that due to its own bulk unaffected by the presence of "roots" projecting into the substratum. This is explained by the fact that volcanic mountains are not produced by compression forcing the crust both downwards and upwards, as other mountain masses are supposed to have been produced, but are mere heaps of materials derived either from the crust or the substratum, and probably drawn from a considerable area. Hence they are balanced not by "roots" projecting immediately below them, but by a slight depression or sagging of the crust over a wide area, and thus having little effect on the rate of the pendulum. In the case of the Falkland Islands, however, the force of gravity is less than it ought to be, and this exception affords an interesting confirmation of
VOL. I. E
the general theory. For these are not volcanic, but are true continental islands, forming the outer margin of the old continent of South America, though now 350 miles from land; and thus, being surrounded by water instead of by much heavier land, the force of gravity is somewhat reduced, water having here replaced a denser mass of land.
The Argument from Volcanic Phenomena.
We now come to the more special researches of Mr. Fisher, which throw so much light on the hitherto unexplained phenomena of volcanoes. By means of some recent experiments on the melting-point and specific heat of rocks, made at his suggestion, he arrives at the conclusion that the average thickness of the earth's crust on lands near the sea-level is only about 18 miles. Its density is estimated at 2·68, water being 1, and the density of the liquid substratum at 2·96.1 With these new data it appears that if the melted substratum were an inert mass it would have cooled at such a rate that the crust would have attained its present thickness in about eight million years. But geologists are almost unanimously of opinion that any such period as this is absurdly too small, and that to account for the phenomena presented by the known series of rocks and their included organic remains, the very least time that must be allowed is one hundred million years. The conclusion Mr. Fisher draws from this discrepancy is, that the substratum is not inert but energetic, that is, that it is in a state of movement or circulation, convection currents continually bringing up fresh heat from below and thus preventing the crust from solidifying so rapidly as if there were no such currents. A cause of such currents is found in the friction produced by tidal action in the liquid mass, which Professor George Darwin has shown to be very great, and to be at a maximum in the central portions.2
Gravity having approximately its normal value all over
1 For these conclusions see the Appendix to Physics of the Earth's Crust.
2 This is pointed out in a paper by Mr. Fisher of a later date than his volume above referred to; in Proc. Cambridge Phil. Soc., 1892.
the globe at the sea-level, it is evident that there must be some denser matter under the oceans to make up for the much less density of the water, which is at least three miles deep on the average. A very refined mathematical investigation shows that this can only be brought about by the sub-oceanic crust being both thinner and denser than under the continents, the denser portion being the upper layer. This distribution of matter may, it is supposed, be due to extensive outflows of heavy basalt over the original depressions forming the ocean floors, at some early period of their history.
The physical constitution of the liquid matter forming the substratum is the next point to be considered, and is one of the highest importance, since it is evidently what determines both volcanic action and a large portion of the disturbances to which the crust is subject. Many geologists are of opinion that the phenomena of volcanic action can only be explained on the supposition that the molten matter forming the interior of the globe holds in solution enormous quantities of water-vapour and other gases; and there is ample evidence that melted lavas and slags do contain such gases, which they give out on becoming solid. Thus Mr. Scrope, in his great work on Volcanoes, says:—
"There unquestionably exists within and below volcanic vents, a body of lava of unknown dimensions, permanently liquid at an intense temperature, and continually traversed by successive volumes of some aeriform fluid, which escape from its surface—thus presenting all the appearance of a liquid in constant ebullition."
And again:—
"If any doubt should suggest itself, whether this fluid is actually generated within the lava, or only rises through it, having its origin in some other manner, it must be dispelled by the evidence afforded in the extremely vesicular or cellular structure of very many erupted lavas, not merely near the surface, but throughout the mass, showing that the aeriform fluid in these cases certainly developed itself interstitially in every part."
Professor Judd, in his volume on the same subject, shows that the presence of these gases in lava is in accordance with Henry's law, that liquids are able to absorb
E 2
gases to an amount proportioned to the pressure they are under, and with the fact that molten substances do actually absorb large quantities of gases. He says:—
"Silver in a state of fusion is able to absorb 22 times its volume of oxygen gas. When the metal is allowed to cool this gas is given off, and if the cooling takes place suddenly a crust is formed on the surface, and the phenomenon known as the spitting of silver is exhibited. Sometimes during this operation miniature cones and lava-streams are formed on the surface of the cooling mass, which present a striking resemblance to those formed on a grand scale on the surface of the globe. The researches of Troost and others have shown that molten iron and steel possess the property of absorbing considerable quantities of oxygen, hydrogen, carbonic acid, and carbonic oxide, and that these gases are given off when either the temperature or the pressure is diminished.… Von Hochstetter has shown that when molten sulphur is exposed to a temperature of 262° Fahrenheit, and a pressure of two or three atmospheres, in the presence of steam, it is found that the sulphur absorbs a considerable quantity of water, which is given off again with great violence from the mass as it undergoes solidification. The hardened crust which forms on the surface of the sulphur is agitated and fissured, miniature cones and lava-streams being formed upon it, which have a striking resemblance to the grander phenomena of the same kind exhibited upon the crust of the globe."
He then goes on to show that the enormous quantity of steam and other gases given off during volcanic action and from flowing lava-streams, can only be accounted for by supposing that the molten rock from which they are derived contains these gases to an amount equal to many times their volume; and that the same fact is indicated by the liquefied gases that are found in the cavities of the crystals of volcanic products which have consolidated under great pressure, such as granites, porphyries, and other rocks of allied nature.
There can, therefore, be no doubt as to the fact of the liquid substratum containing in its substance an enormous quantity of gases, the principal being water-vapour, but how the gases came there is less certain; nor does it materially concern us. Some think that these gases have been largely derived from sea-water, which has found its way by percolation to the heated interior; but there are
1 International Scientific Series, vol. xxxv., "Volcanoes," p. 355,
many difficulties in this view. Others, with whom is Mr. Fisher, think that they form an essential constituent of the primeval globe, and that, instead of being derived from the ocean, it is more probable that the ocean itself has been derived from the vapours which have been always escaping from the interior. Leaving this question as one of comparatively little importance for the present discussion, we have now to point out how the facts, that the fluid substratum is saturated with water-vapour and other gases, and is also subject to convection-currents continually bringing superheated matter up to the lower surface of the crust, enable us to explain the special difficulties as to volcanic eruptions alluded to in the early portion of this chapter.
The first of these difficulties is, that neighbouring volcanoes of very different heights act quite independently, a fact which is supposed to be inconsistent with the idea that both are in connection with the same molten interior. It seems, however, to have been assumed that a mere fissure or other aperture extending from the surface to the substratum, or from the substratum to the surface, would necessarily be followed by an outflow of lava, even though the opening terminated at the summit of a mountain many thousand feet above the sea level. But it is evident that on the theory of a molten interior, with a crust of somewhat less specific gravity resting upon it in hydrostatic equilibrium, nothing of the kind would happen. When a hole is bored through an extensive ice-field, whether on a lake or in the Arctic Ocean, the water does not spout up through the aperture, but merely rises to the same level as it would reach on the sides of a detached block of floating ice, or on the outer margin of the ice-field itself. The facts that the fluid on which the crust of the earth rests is intensely heated, and that the crust is continuous over its whole surface, can make no difference in the behaviour of the fluid and the solid, so as to cause the molten rock to rise with great violence thousands of feet above its mean level whenever an aperture is made; and this is the more certain when we take account of the fact, which may now be considered to be well established,
that the crust floats on the fluid interior, and that it is so thin and weak, comparatively speaking, that it cannot resist a strain equal to its own weight, but must bend or fracture so as to keep every part in approximate hydrostatic equilibrium. Volcanic action, especially continuous and permanent volcanic action like that of Stromboli and Kilauea, cannot, therefore, be explained by the mere existence of a thin crust and a molten interior; but it is well explained by the presence in the molten mass of vast quantities of gases existing under enormous pressure, and ready to escape with tremendous force whenever that pressure is greatly diminished, and the molten material that contains it lowered in temperature.
Let us now endeavour to trace what will happen when a fissure is opened gradually from below upwards till it reaches the surface. Owing to hydrostatic pressure the fluid will rise in the fissure, and in doing so will be subject to some cooling and diminution of density, which, as we have seen, will lead to a liberation of some of the contained gas. The pressure of this gas will aid in extending the fissure, and the liquid will continue to rise till it reaches the level of hydrostatic equilibrium, which would be somewhere about two miles below the surface. But throughout the whole mass of the liquid in the fissure, and for some depth below the under surface of the crust, there would be a continual liberation of intensely heated gases. These would no doubt carry with them in their upward rush a portion of the liquid matter which had risen from below, but they would also, owing to their intensely heated condition, melt off some portions of the rocky walls of the fissure, and thus give to the ejected volcanic products a local character. We here see the explanation of the supposed difficulty of the individuality of neighbouring volcanoes and the diversity of their products, and also of the fact of an eruption of lava from the crater of a lofty mountain while the liquid lava of one close by, and thousands of feet lower, maintains its usual level. Kilauea we may suppose to owe its permanently molten lake to a siphon-like passage through which a constant flow of heated gases is maintained, and which
suffices to keep its lava in permanent ebullition; while the lofty Mauna Loa has its vent usually blocked up, and may owe its occasional eruptions to an accumulation of gases in some deep-seated cavities which, at long intervals, become sufficiently powerful to burst away the obstacle and pour out a quantity of melted material derived from the sides of the channels through which they make their way upward.
The phenomena presented by the crater of Kilauea, where an extensive lava-lake remains in a constant state of ebullition while keeping approximately the same level, can only be explained by the upward percolation of heated gases in moderate and tolerably uniform streams, sufficient to keep up the melting temperature of the lava; while occasional more powerful outbursts throw up jets or waves of the molten matter, or sometimes break up the crust that has formed over portions of the lake. Here, evidently, there is no eruption in the ordinary sense, no fresh matter is being brought up from below, but only fresh supplies of intensely heated gases sufficient to keep the lava permanently liquid, and to produce the jets, waves, and fountains of lava, and the strange surging, swirling, and wallowing motions of the molten mass, so well described by Miss Bird, Lord George Campbell, and other competent observers.
The sketch now given of Mr. Fisher's investigations as to the nature of the molten interior of the earth and of the crust which overlays it, only covers a small portion of the ground traversed in his work. He there deals also with the more difficult questions of the stresses produced by the contraction of the cooling earth, and the various theories that have been suggested to explain the great inequalities of its surface. The origin of the great oceanic depressions and of the vast mountain masses that everywhere diversify the continental areas, and the causes that have produced the compression, upheaval, folding and crumpling of the rocks at every period of geological history, are all discussed, and some light is thrown upon these confessedly obscure and very difficult problems.
But whatever doubts may still exist as to the exact causes of these last-named phenomena do not apply to those to which the present chapter is mainly devoted. So many distinct but converging lines of evidence indicate the existence of a molten substratum holding in solution, in accordance with well-known physical laws, great quantities of steam and other gases, and show that the crust covering it is a very thin one—while the hypothesis of such a substratum and thin floating crust so well explains the curious phenomena of great masses of strata thousands of feet thick, yet from top to bottom bearing indications of having been deposited in shallow water, and the no less singular fact of a corresponding recent subsidence in all great river-deltas, and also clears up so many difficulties in the modes of volcanic action and the diversity of volcanic products— that we can hardly doubt the correctness of the hypothesis. And though at first sight the idea of our being separated by a thickness of only eighteen miles of rock from a layer of molten lava of unknown depth may appear somewhat alarming, yet the very tenuity and fragility of the crust may itself be a source both of safety and of utility. While sufficiently thick to secure us from any injurious or even perceptible effects of internal heat, except in volcanic or earthquake areas, it yet gives us the possibility and even the promise of an inexhaustible source of heat and power at such a moderate distance that we may some day be able to utilise it. On the other hand, the thin crust so readily and constantly adjusts itself to all the alternations of strain and pressure to which it may be exposed, that we are thereby secured from the occurrence of vast cataclysms capable of endangering the existence of any considerable portion of our race. A solid earth might, possibly, not be so safe and stable as is Our Molten Globe.
APPENDIX
IN an interesting article (Science, June and July, 1899) Mr. T. C. Chamberlin has discussed the theory of the formation of our earth by meteoric accretion instead of by the condensation of a gaseous or liquid ring left behind by the contracting sun; and he suggests that the process of accretion might have been so slow that the heat produced would never have been sufficient to produce a liquid globe, and that the somewhat irregular mass of matter partially solid or in parts liquefied, would better explain many geological phenomena and might lengthen indefinitely the period of cooling, so as to obviate the difficulty as to a too limited period of solidity given by the calculations of Lord Kelvin.
As the meteoritic theory of the universe is now becoming widely accepted as being in accordance with a vast body of facts, and offering none of the almost insuperable difficulties presented by the nebular hypothesis, it may be thought that it contradicts the theory of a liquid interior so strongly supported by the observations and calculations summarized in the present chapter. This, however, is by no means necessarily the case. It may be that the rate of meteoric accretion has greatly varied. At first it may have been so slow, and the impact of the constituent meteors may have been so comparatively weak owing to the slight force of their mutual attraction, that the nascent globe long remained solid. But when it had grown to a considerable bulk its attractive force would render the impact of the falling meteors so great as to produce partial liquidity. If at this stage the larger proportion of the contents of the meteoric ring were drawn in to the growing mass, and especially if several large aggregations of matter successively fell in, the outer portions of the globe might become liquid while the interior remained solid, though imperfectly aggregated, and with numerous hollows filled with gases or liquids. It may, therefore, be the case that under the thin crust
indicated by the facts adduced by Mr. Fisher there exists a molten interior of unknown thickness, although below this molten matter there may be a core, perhaps of equal bulk, formed mainly of solid but imperfectly aggregated matter, permeated by liquids and gases. Whether such a mixed constitution of our globe accords with the evidence of the pendulum experiments must be left to the mathematicians to determine.
CHAPTER IV
THE ICE AGE AND ITS WORK
I. ERRATIC BLOCKS AND ICE-SHEETS
IT is little more than fifty years ago that one of the most potent agents in modifying the surface-features of our country was first recognised. Before 1840, when Agassiz accompanied Buckland to Scotland, the Lake District, and Wales, discovering everywhere the same indications of the former presence of glaciers as are to be found so abundantly in Switzerland, no geologist had conceived the possibility of a recent glacial epoch in the temperate portion of the northern hemisphere. From that year, however, a new science came into existence, and it was recognised that only by a careful study of existing glaciers, of the nature of the work they now do, and of the indications of the work they have done in past ages, could we explain many curious phenomena that have hitherto been vaguely regarded as indications of diluvial agency. One of the first-fruits of the new science was the conversion of the author of Reliquiæ Diluvianæ—Dr. Buckland, who, having studied the work of glaciers in Switzerland in company with Agassiz, became convinced that numerous phenomena he had observed in this country could only be due to the very same causes. In November, 1840, he read a paper before the Geological Society on the "Evidences of Glaciers in Scotland and the North of England," and from that time to the present the study of glaciers and of their work has been systematically pursued with a large amount of success.
One after another, crude theories have been abandoned, facts have steadily accumulated, and their logical though cautious interpretation has led to a considerable body of well-supported inductions on which the new science is becoming firmly established. Some of the most important and far-reaching of these inductions are, however, still denied by writers who have a wide acquaintance with modern glaciers; and as several works have recently appeared on both sides of the controversy, the time seems appropriate for a popular sketch of the progress of the glacial theory, together with a more detailed discussion of some of the most disputed points as to which it seems to the present writer that sound reasoning is even more required than the further accumulation of facts.1
In the last century, Swedenborg, Linnæus, Pallas, De Luc, and many other eminent writers took notice of the remarkable fact that in Scandinavia, Russia, Germany, and Switzerland detached rocks or boulders were found, often in great abundance and of immense size, and of a kind that did not exist in situ in the same district, but which were often only to be discovered in remote localities, sometimes hundreds of miles away. Those who ventured to speculate on the origin of these travelled rocks usually had recourse to water-power to account for their removal; and as their large size and often elevated position required some unusual force to carry them, there arose the idea of enormous floods sweeping over whole continents; and for a long time this diluvial theory was the only one that appeared to be available, although the difficulties of its application to explain all the phenomena became greater the more closely those phenomena were studied. Still, there was apparently no other known or conceivable means of accounting for them, and for the enormous mounds of
1 The works referred to are:—Do Glaciers Excavate? by Prof. T. G. Bonney, F.R.S. (The Geographical Journal, vol. i., No. 6); The Glacial Nightmare and the Flood, by Sir H. H. Howorth, M.P., F.R.S.; Fragments of Earth Lore, by Prof. James Geikie, F.R.S.; Man and the Glacial Period, by Prof. G. F. Wright, F.G.S.A.; La Période Glaciaire, by A. Falsan; and the Glacialists' Magazine, edited by Percy F. Kendall, F.G.S.; from which works, and from those of Lyell, Ramsay, Geikie, and the American geologists, most of the fact referred to in these chapters are derived.
gravel or clay intermixed with boulders which often accompanied them; and the efforts of geologists were therefore directed to the discovery of how the water-power had acted, and by what means the supposed floods could have been produced.
But there were not wanting men who saw that no action of water alone could account for the facts. Sir James Hall pointed this out with regard to erratics on the Jura, whose source was undoubtedly in the far distant Alps; and Mr. Grainger, in America, described some of the parallel grooves and flutings running for nearly a mile in Ohio, strongly arguing that no action of running water could have produced them, but that an agent was required the direction of whose movement was fixed and unalterable for long distances and for a great length of time. No light was, however, thrown on the problem till 1822, when Venetz, a Swiss engineer, finding that existing glaciers varied in extent from year to year and that historical records showed them to have considerably increased during the last eight centuries, was further led to observe that, long before the historical era the glaciers had been immensely more extensive, as shown by the smooth and rounded rocks, by longitudinal scratches and grooves pointing down the valleys, and by numbers of old moraines exactly similar in form and materials to those deposited by existing glaciers. He read a paper before the Helvetic Society of Natural History, and urged that glaciers once stretched down the Rhone valley as far as the Jura, and there deposited the erratic blocks which had so puzzled the diluvialists to explain.
Other writers soon followed the clue thus given. In 1835 Charpentier, after a close study of the erratic blocks and of their sources, adopted the views of Venetz. Agassiz followed, and by his strenuous advocacy did much to spread correct views as to the former extension of the Alpine glaciers, and their capability of explaining the numerous superficial phenomena which in all northern countries had been thought to afford proofs of enormous floods and of the submergence of a large part of Europe under a deep sea. He has, therefore, gained the reputation of being the
originator of the modern school of glacialists, which undoubtedly owes much to his energy, research, and powers of exposition, though all the more important facts, as well as the logical conclusions to be drawn from them, had been pointed out by previous writers.
Before proceeding further, it will be well to give a brief outline of the phenomena which lead to the conclusion that glaciers have formerly existed in districts and countries where even perpetual snow on the mountain tops is now unknown. These may be briefly classed as: (1) Moraines and drifts; (2) Rounded, smoothed, or planed rocks; (3) Striæ, grooves, and furrows on rock-surfaces; (4) Erratics and perched blocks.
(1) Moraines are those heaps or ridges of rock and other débris which are deposited on the surface of a glacier from the precipices or mountain slopes which border it, and which form what are termed lateral and medial moraines while upon it, and terminal moraines when, being gradually discharged at its end, either from above or from beneath it, they form great heaps of rock and gravel corresponding in outline and extent to that of the terminal ice-cliff. Such moraines can be seen on and near all existing glaciers, and their mode of formation and characteristics are perfectly well known. If the glacier is continuously retreating, then the terminal moraine will form more or less irregular heaps over the surface the glacier has formerly covered; but when, as is usually the case, the glacier remains stationary for a considerable period, then the terminal moraine will have a definite form, and will often stretch quite across the valley, but presenting one or more openings through which the glacier stream has cub its way. Such moraines form steep mounds, usually curved and often very regular, seeming from a little distance to block up the valley like an artificial earthwork. Among hundreds that might be enumerated good examples may be seen in Glen Isla (Forfarshire), in the Troutbeck valley near Windermere, and in Cwm Glas, on the north side of Snowdon, this latter being so regularly curved, evenly sloped, and level-topped as to look from below exactly like an ancient fortification. The char-
acteristic features of moraines are, their position in valleys where there are other indications of glacial action, their steep slopes and often level tops, but especially their composition of earth, stones, and gravel, with large fragments of rock irregularly scattered through them from top to bottom without any sign of stratification, while usually one or more large blocks rest upon their summits in positions where they could only have been left by the retreat of the glacier, or possibly stranded from floating ice. The accompanying cut (Fig. 9) shows the surface of part of the

FIG 9.—SURFACE OF PART OF GREAT TERMINAL MORAINE.
great terminal moraine of the North American ice-sheet in south-eastern Massachusetts, from the seventh Report of the United States Geological Survey. It presents a more barren and stony surface than most of our moraines, which are usually grass-covered. Fig. 10 is a view of Easedale Tarn above Grasmere, showing a series of moraine heaps beyond the lake. Where extensive glaciers have covered large areas of nearly level ground the moraines form great sheets extending for many miles, often concealing the original contours of the country, and then receive the general name of drift. The composition
of drift is usually the same as that of well-marked moraines, large blocks of stone being distributed throughout its mass. It is this which mainly distinguishes drift from alluvial or shore deposits, in which the materials are always more or less assorted and stratified; but the angular forms of many of the contained blocks and the striated surfaces of others are also characteristic. Besides the terminal moraines of extinct glaciers, lateral moraines are also left along the

FlG. 10.—EASEDALE TARN, WITH, MORAINES.
slopes of open valleys from which glaciers have retreated. As a whole, moraines are well distinguished from all accumulations formed by water, and it has not been shown that any other agency than glaciers is capable of forming them. In all recently glaciated countries they are to be found more or less frequently, and thus afford an excellent first indication of the former existence of glaciers.
(2) Smoothed and rounded rocks, called in Switzerland "roches moutonnées," from their supposed resemblance at
a distance to sheep lying down, are perhaps the most general of all the indications of glacial action. Every glacier carries with it, embedded in its under surface, numbers of rocks and stones, which, during the slow but unceasing motion over its bed, crush and grind down all rocky projections, producing in the end gently rounded or almost flat surfaces even on the hardest and toughest rocks. In many of the valleys of Wales, the Lake District, and Scotland every exposed rock has acquired this characteristic outline, and the same feature can be traced on all the rocky slopes, and often on the summits of the lesser heights; and the explanation of how these forms have been produced is not a theory only, but has been observed in actual operation in the accessible portions of many glaciers. Rocks and stones are to be seen embedded in the ice and actually scratching, grooving, and grinding the rock beneath in their slow but irresistible onward motion. The rocky islets in Windermere, Ullswater, and other lakes, as well as the Thousand Islands of the St. Lawrence, are thus ice-ground; and the amount of the grinding can often be seen to be proportional to the pressure and motion of the advancing glacier. I recently noticed in the marshy alluvial plain above Derwentwater a projecting rock which has been ground down to so regular a curve as to look like a portion of an enormous globe buried in the earth. By rough measurement and estimate this rock was about 250 feet across, and 20 or 30 feet high. It was formed of hard slate, with numerous quartzite veins, the whole ground down to a uniform spherical surface. It had evidently once been an island in the lake, having a much broader base now hidden by the alluvium, and may originally have been one of those abrupt craggy rocks a few hundred feet high, which, owing to their superior hardness or tenacity, resisted ordinary denudation, and which, when above the old ice-level, form those numerous "pikes" which add so much to the wild and picturesque scenery of the district. Looking at such rocks as this, with outlines so utterly unlike any that are produced in similar formations by sub-aerial denudation—and they are to be seen by scores in all glaciated regions—we cannot but conclude
VOL. I. F
that the ice-tool has done more than merely rub off the angles and minor prominences, and that it has really ground away rocky hills to an unknown but very considerable extent; and this conclusion is, as we shall see, supported by a very large amount of confirmatory evidence. It may be noted that ice-ground rocks usually show the direction in which the ice has moved, by the side opposed to the motion being more completely smoothed than the lee side, which often retains some of its ruggedness, having been protected partly by the ice overriding it and partly by
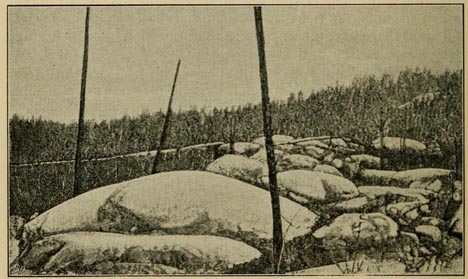
FIG. 11.—ROCHES MOUTONNÉES, IN IDAHO, U.S.A.
the accumulation of its own débris. Where such rocks occur in the higher parts of valleys the smooth side always looks up the valley from which the glacier has descended. In the more open parts of valleys, or in high coombs or cirques, where two or more small ravines meet and where the ice may have been embayed and have acquired a somewhat rotary motion, the rocks are seen to be ground down on all sides into smooth mammillated mounds or hummocks, showing that the ice has been forced into all the irregularities of the surface. An example on a small scale is to be seen in Cwm Glas, on the north side of Snowdon,
above the fine moraine already mentioned, and in many other places around the same mountain. On the whole, considering their abundance in all glaciated regions, and the amount of information they give as to the direction and grinding power of ice, these rounded rocks afford one of the most instructive indications of the former presence of glaciers; and we must also agree with the conclusion of Darwin (in a paper written after studying the phenomena of ice action in North Wales, and while fresh from his

FIG. 12.—STRIATED ROCK-SURFACE AT WASDALE CRAG.
observations of glaciers and icebergs in the southern hemisphere) that "one of the best criterions between the effects produced by the passage of glaciers and of icebergs is boss or dome-shaped rocks." (Fig. 11.) from the United States Geological Survey Reports, is a fair illustration of roches moutonnées.
(3) Striated, grooved, and fluted rocks, though closely connected with the preceding, form a distinct kind of evidence of the greatest value. Most of the bosses of
F 2
rock just described have been exposed to the action of the atmosphere, perhaps since the ice left them, and have thus become more or less roughened or even disintegrated; but where the rocks have been protected by a covering of drift, or even of turf, and have been recently exposed, they often exhibit numerous parallel striæ, varying from the finest scratches to deep furrows a foot or more in diameter. Fine examples are to be seen near the lakes of Llanberis, and they occur more or less frequently in every glaciated country. The accompanying illustration (Fig. 12), which I owe to the kindness of Mr. Godfrey Bingley, of Leeds, shows a striated surface of Shap granite at Wasdale Crag, Westmoreland. Perhaps none of the effects of ice so clearly demonstrate the action of glaciers as opposed to that of icebergs, owing to the general constancy of the direction of the striæ, and the long distances they may be traced up and down slopes, with a steadiness of motion and evenness of cutting power which no floating mass could possibly exert. Sir A. Geikie tells us that in Gareloch, Bute, and Cantyre the striations on the rocks run up and over the ridges, and are as clearly shown on the hill-tops as in the valleys. Mr. D. Mackintosh states (in his paper on the "Ice-sheet of the Lake District and of North Wales") that in the valley above Windermere the striæ cross Rydal Fell, Loughrigg Fell, and Orrest Head, ascending and descending their slopes, often obliquely. But it is in the United States that the most remarkable rock-groovings are to be found, extending over a large portion of the North-eastern States. In his report on "The Rock-scorings of the Great Ice Invasions," Mr. T. C. Chamberlin gives many fine illustrations, from photographs, showing striæ and grooves along sloping, curved, or vertical surfaces, the striæ following the changes of curve, so that the grinding material must have been slowly forced into close contact with the irregular surface. Of one of these examples Mr. Chamberlin says:—
"The climax of adaptability is reached in the striation of warped and twisted surfaces, and of tortuous valleys. One of the most remarkable known instances of this within the limits of photographic illustration is furnished by the great glacial grooves at Kelly's
Island. These exhibit not only the pliancy of the ice, but at the same time its strong hold upon the armature with which it did its work of abrasion, grooving, and striation. For, while these grooves can scarcely be supposed to have been originated de novo by the gouging action of the ice, they are, nevertheless, ploughed with deep furrows, the symmetry, continuity, and peculiar form of some of which are only intelligible on the supposition that they were cut by a single graving tool, held with sufficient tenacity by the ice to execute by a single movement a deep, sharply-defined groove. There is, perhaps, no finer illustration of the pliancy with which the ice yielded to its encompassing barriers, the tenacity with which it held its armature, and withal the pressure that both forced it into compliance with its tortuous channel, and pressed it relentlessly forward."1
Kelly's Island is at the western end of Lake Erie, and in the direction of the striæ to the north-east there is no high ground for about 400 miles. Looking at these facts, I cannot give any weight to the opinions of these who, from observations of existing glaciers, declare positively that ice cannot go up-hill, and can exert no grinding power on level ground.
The two photo-plates here given show, however, that we possess equally fine illustrations of ice-grooving in our own country. Fig. 13, kindly given me by Mr. Percy F. Kendall, well shows the size of the grooves by comparison with the boy's body. It was taken at Barmouth, N. Wales. The other (Fig. 14), I believe at the same place, shows how the ice has been forced into curves by the general form of the rock surface, a phenomenon that indicates a slow-moving plastic material such as glacier-ice is proved to be, and which could not have been caused by any other known agency. The highly polished surface of the rock is well shown in the photograph, though in a less degree in the reproduction.
Erratics and Perched Blocks.
(4) Erratic blocks were among the phenomena that first attracted the attention of men of science. Large masses of granite and hard metamorphic rock, which can be
1 Seventh Annual Report of the United States Geological Survey, p. 179 (Fig. 17). Arrangements have now been made for the preservation of these remarkable examples of icework.
traced to Scandinavia, are found scattered over the plains of Denmark, Prussia, and Northern Germany, where they rest either on drift or on quite different formations of the Secondary or Tertiary periods. One of these blocks, estimated at 1,500 tons weight, lay in a marshy plain near St. Petersburg, and a portion of it was used for the pedestal of the statue of Peter the Great. In parts of North Germany they are so abundant as to hide the

FIG. 13.—ROCK-GROOVINGS AT BARMOUTH.
surface of the ground, being piled up in irregular masses forming hills of granite boulders, which are often covered with forests of pine, birch, and juniper. Far south, at Fürstenwalde south-east of Berlin, there was a huge block of Swedish red granite, from one half of which the gigantic basin was wrought which stands before the New Museum in that city. In Holstein there is a block of granite 20 feet in diameter; and it was noticed by
De Luc that the largest blocks were often found at the greatest distance from the parent rock, and that this fact was conclusive against their having been brought to their present position by the action of floods.
Erratic Blocks in Switzerland.
It is, however, in Switzerland that we find erratic blocks which furnish us with the most conclusive testi-

FIG. 14.—ROCK-GROOVINGS NEAR BARMOUTH.
mony to the former enormous extension of glaciers; and as these have been examined with the greatest care, and the facts, as well as the main inductions from the facts, are generally admitted by all modern writers, it will be well to consider them somewhat in detail. It will be found that they give us most valuable information both as to the depth and extension of ancient glaciers, and also as to the possibilities of motion in wide-spreading ice-sheets.
The most important of these facts relate to the erratic blocks from the higher Alps, which are found on the flanks of the Jura Mountains wholly formed of limestone, on which it is therefore easy to recognise the granites, slates, and old metamorphic rocks of the Alpine chain. These erratic blocks extend along the Jura range for a distance of 100 miles, and up to a height of 2,015 feet above the Lake of Neufchatel. The first important point to notice is, that this highest elevation is attained at a spot exactly opposite, and in the same direction as, the Rhone valley, between Martigny and the head of the Lake of Geneva, while north or south of this point they gradually decline in elevation to about 500 feet above the lake. The blocks at the highest elevation and central point can be traced to the eastern shoulder of Mont Blanc. All those to the south-west come from the left-hand side of the lower Rhone valley, while those to the north-east are all from the left side of the upper Rhone valley and its tributaries. Other rocks coming from the right-hand side of the upper Rhone valley are found on the right-hand or Bernese side of the great valley between the Jura and the Bernese Alps.1
Now, this peculiar and definite distribution, which has been worked out with the greatest care by numerous Swiss geologists, is a necessary consequence of well-known laws of glacier motion. The débris from the two sides of the main valley form lateral moraines which, however much the glacier may afterwards be contracted or spread out, keep their relative position unchanged. Each important tributary glacier brings in other lateral moraines, and thus when the combined glacier ultimately spreads out in a great lowland valley the several moraines will also spread out, while keeping their relative position, and never crossing over to mingle with each other. So soon as this definite position of the erratics was worked out it became evident that the first explanation—by a great submergence during which the lower Swiss valleys were arms of the sea and the Rhone glacier broke off in icebergs which carried the erratics across to the Jura—was
1 See Map at p. 128.
altogether untenable, and that the original explanation of Venetz and Charpentier was the true one. Sir Charles Lyell, who had first adopted the iceberg theory, gave it up on examining the country in 1857 and ascertaining that the facts were correctly stated by the Swiss geologists; and there is at the present day no writer of the least importance who denies this. Sir Henry Howorth, who is one of the strongest opponents of what he considers the extreme views of modern glacialists, gives a full summary of the facts as to the old Rhone glacier from Charpentier. He states that between Martigny and St. Maurice the moraine débris on each side of the valley shows the glacier to have reached a height of 3,000 feet above the river; farther on, where the valley widens over the Lake of Geneva, it sank to 2,600 feet, while on the Jura itself it seems to have been again raised to 3,000 feet at its highest point1; and he quotes Charpentier's general conclusion:—
"It goes without saying that not only all the valleys of the Valais were filled with ice up to a certain height, but that all lower Switzerland in which we find the erratic débris of the Rhone valley must have been covered by the same glacier. Consequently all the country between the Alps and the Jura, and between the environs of Geneva and those of Soleure, has been the bed of a glacier."
And then, after quoting the observations of Agassiz on the same phenomena and of those of North America, he gives his own conclusions in the following words:—
"It is plain to those who would look without prejudice that the rounded and mammilated surfaces, the scratched, polished, and grooved rocks, and a great number of the phenomena which accompanied the distribution of the boulders and the drift, are consistent only with the fact that in the last geological age there was an immense development of glaciers which occupied not only the high ranges of the Alps and the Dovrefjeld, but the secondary ranges and lower heights of the continents of Europe and North America. This conclusion seems supported by every form of converging evidence, and is apparently beyond the reach of cavil. So far there is no question at issue."2
1 These figures are almost certainly incorrect, as the upper surface of the glacier must have had a considerable downward slope to produce motion. The recent work of M. Falsan, La Période Glaciaire, gives the thickness as about 3,800 feet at the head of the lake and 3,250 feet at Geneva.
2 The Glacial Nightmare and the Flood, p. 208.
We may take it, therefore, that the views of Charpentier, Agassiz, and Sir Charles Lyell as to the extent and thickness of the great Rhone glacier are admitted to be correct, or, at least, not to be exaggerated, by the most strenuous opponents of the extreme glacialists. We may, therefore, use this as a fixed datum in our further investigations, and I think it will be found to lead us irresistibly to conclusions which in other cases these writers declare to be inadmissible.
Erratic Blocks in North America.
We must now consider briefly the distribution of erratics in North America, because they present some peculiar features and teach us much concerning the possibilities of glacier motion.
An immense area of the North-eastern States, extending south to New York, and then westward in an irregular line to Cincinnati and St. Louis, is almost wholly covered with a deposit of drift material, in which rocks of various sizes are embedded, while other rocks, often of enormous size, lie upon the surface. These blocks have been carefully studied by the American geologists, and they present us with some very interesting facts. Not only are the distances from which they have been transported very great, but in many cases they are found at a greater elevation than the place from which they must have come. Professor G. F. Wright found an enormous accumulation of boulders on a sandstone plateau in Monroe County, Pennsylvania. Many of these boulders were of granite, and must have come either from the Adirondack Mountains 200 miles to the north, or from the Canadian Highlands still farther away. This accumulation of boulders was 70 or 80 feet high, and it extended many miles, descending into a deep valley 1,000 feet below the plateau in a nearly continuous line forming part of the southern moraine of the great American ice-sheet.
On the Kentucky hills, about twelve miles south of Cincinnati, conglomerate boulders containing pebbles of red jasper can be traced to a limited outcrop of the same rock
in Canada to the north of Lake Huron, more than 600 miles distant, and similar boulders have been found at intervals over the whole intervening country. In both these cases the blocks must have passed over intervening valleys and hills, the latter as high or nearly as high as the source from whence the rocks were derived. Even more remarkable are numerous boulders of Helderberg limestone on the summit of the Blue Ridge in Pennsylvania, which must have been brought from ledges at least 500 feet lower than the places upon which they now lie. The Blue Ridge itself shows remarkable signs of glacial abrasion, in a well-defined shoulder marking the southern limit of the ice (as indicated also by heaps of drift and erratics), so that Mr. Wright concludes that several hundred feet of the ridge have been worn away by the ice.
The crowning example of boulder transportation is, however, afforded by the blocks of light grey gneiss discovered by Professor Hitchcock on the summit of Mount Washington, over 6,000 feet above sea-level, and identified with Bethlehem gneiss, whose nearest outcrop is in Jefferson, several miles to the north-west, and 3,000 or 4,000 feet lower than Mount Washington.
These varied phenomena of erratic blocks and rock striations, together with the enormous quantity of boulder-clay and glacial drift spread over the whole of the Eastern States, terminating southward in a more or less abrupt line of mounds having all the characteristics of an enormous moraine, have led American geologists to certain definite conclusions in which they all practically agree. It may be well first to give a notion of the enormous amount of the glacial débris under which a large part of the Eastern States is buried. In New England these deposits are of less thickness than farther south, averaging from 10 to 20 feet over the whole area. In Pennsylvania and New York east of the Alleghanies, the deposits are very irregular, often 60 or 70 feet thick and sometimes more. West of the Alleghanies, in New York, Pennsylvania and Ohio, the thickness is much greater, being often 150 or 200 feet in the wide valleys and 40 or 50 feet on many of the uplands. Professor Newberry calculates that in
Ohio it averages 60 feet deep over an area of 25,000 square miles.
The North-American Ice-sheet.
The direction of the striæ and of the travelled boulders together with the form of the great terminal moraines show that there must have been two main centres of outflow for the ice-sheet, one over Labrador, the other over the Laurentian Highlands north of Lake Superior. The southern margin of the drift may be roughly represented by portions of circles drawn from these two points as centres. The erratics on the summit of Mount Washington show that the ice-sheet must have been a mile thick in its neighbourhood, and much thicker at the centres of dispersion, while the masses of drift and erratics on plateaus 2,000 feet high near its southern boundary indicate a great thickness at the termination. The Laurentian plateau is now about 2,000 feet above the sea-level, but there are numerous indications from buried river channels, filled with drift and far below the sea, which lead to the conclusion that during the Ice Age the land was much higher. That snow can accumulate to an enormous extent over land of moderate height when the conditions are favourable for such an accumulation is shown by the case of Greenland, the greater part of whose surface is a vast plateau of ice flowing outward by numerous glaciers into the sea. The centre of this plateau where Dr. Nansen crossed it was over 9,000 feet above sea-level, and it may be very much higher farther north. It, therefore, seems probable that the great American ice-sheet was, at least, as high and perhaps much higher, and this would give sufficient slope for the flow to the southern border. Of course, during the successive stages of the glaciation there may have been numerous local centres from which glaciers radiated, and during the passing away of the Ice Age these local glaciers would have left striæ and other indications of their presence. But so much of the area covered by the drift—all, in fact, south of the New England mountains and the Great Lakes—is undulating ground, hill, valley, and plateau of moderate height, that here all the
phenomena seem to be due to the great confluent ice-sheet during the various phases of its advance and its passing away.
Sir Henry Howorth, in his very instructive work already quoted, denies the existence and even the possibility of such ice-sheets as those here indicated as having occurred in North America and Europe. He maintains that ice of the requisite thickness could not exist, as it would be crushed or liquefied by its own weight; and further, that if it existed it could not possibly move over hundreds of miles of generally level country, passing over hills and valleys and carrying with it, either on its surface or in its lower strata, the enormous quantity of boulders, gravel, and clay which we find everywhere overlying the present surface of the ground. No doubt the difficulty does seem an enormous one, but I think that it can be shown to be not so great as it seems; and it is certainly by no means so insuperable as that of the apocryphal floods, or "waves of translation" as they have been called, to which he imputes the phenomena. He asks us to believe in one or more gigantic waves sweeping over Eastern North America, carrying boulders to the summit of Mount Washington, nearly 6,000 feet high, scattering others over an area which is roughly 1,000 miles from east to west and 600 from north to south, and in its course producing those wonderful striæ, grooves, and furrows in the rocks photographed in the American reports, and the enormous extent of smoothed and rounded rock surfaces that is found over this wide area.
But, besides these, there are two other phenomena absolutely inconsistent with a diluvial agency. One is the enormous deposits of fine compact clay bearing rounded and scratched stones thickly scattered through it, utterly unlike any deposit produced by water, which would necessarily leave the stones hundreds of miles behind the place to which the fine mud would be carried. The other is the existence of well-defined heaps, mounds, and ridges of gravel and boulders, forming the terminal moraine of the ice-sheet. This is exactly similar in general form and structure to the moraines left by the old Alpine or North
British glaciers, and if the former could have been produced by a flood so could the latter. But the American terminal moraine runs across the country almost irrespective of its contour, and is often as well marked on plateaus as in valleys and on the intermediate slopes. Moreover, this moraine often lies on the southern slope of the hills draining towards the Mississippi valley; and we are asked to believe that a flood vast enough to carry gravel and rocks for hundreds of miles to such a position, left them all stranded on a slope down which it must have been rushing with increased velocity and without hindrance towards the Gulf of Mexico! So far as I know, Sir Henry Howorth is absolutely alone among living writers in his diluvial theories, and I only give this brief statement of their overwhelming impossibilities because his book is so interesting, and his assertions that his theory explains all the facts are so confident and so often repeated, that they are likely to confuse the judgment of readers who have not paid special attention to the subject.
The Rhone Glacier and Ice-sheet.
Returning to the main question, of the possibility of glaciers or ice-sheets moving over long distances of generally level ground with intervening hills and valleys, there is an important piece of evidence, the bearing of which appears to have been overlooked by objectors. The former existence of the great Rhone glacier depositing erratics on the slopes of the Jura from beyond Geneva on the southwest to Soleure on the north-east, is universally admitted. This glacier passed out of the gorge between the Dent du Midi and the Dent de Morcles, and a little below St. Maurice entered on the alluvial plain which extends to the lake. From this point to Geneva, a distance of about 60 miles, may be considered a level plain, the descent into the lake being balanced by the ascent out of it. Yet it is admitted that the glacier did move over this distance, since erratics which can be traced to their source on the left of the valley below Martigny are found near that city. But the main part of the glacier curved
round to the right across the Lake of Neufchatel, and extended at leastas far as Soleure, a distance of about 90 miles. (See Map at page 128.) To do this it must have ascended 500 or 600 feet to the country around Fribourg, and before reaching Soleure must have passed over a hill 300 or 400 feet higher. Yet on the flanks of the Jura above Soleure there are erratics which have been carried on the surface of the glacier from the east side of the valley below Martigny; and close to Soleure itself there are remains of a terminal sub-glacial moraine of compact boulder-clay. Sir Charles Lyell describes this as—
"an unstratified mass of clay or mud, through which a variety of angular and rubbed stones were scattered, and a marked proportion of the whole were polished and scratched, and the clay rendered so compact, as if by the incumbent pressure of a great mass of ice, that it has been found necessary to blow it up with gunpowder in making railway cuttings through part of it. A marble rock, of the age of our Portland stone, on which this old moraine rests has its surface polished like a looking-glass, displaying beautiful sections of fossil shells, while occasionally, besides finer striæ, there are deep rectilinear grooves, agreeing in direction with the course in which the extinct glacier moved according to the theory of M. Guyot before explained." 1
It is evident that, to have produced such effects as are here described, the glacier must have extended much beyond Soleure, and have been very thick even there. It thus proves to demonstration that a glacier can travel for 100 miles over a generally level country, that it can pass over hills and valleys, and that, even near its termination, it can groove, and grind, and polish rocks, and deposit large masses of hard boulder-clay. And all this was done by a single glacier issuing from a comparatively narrow valley, and then spreading out over an area many times greater than that of its whole previous course. In this case it is clear that such a vast mass of ice, constituting a veritable ice-sheet on a small scale, could not have derived its motion solely from the push given to it by the parent glacier at St. Maurice. Neither could gravitation derived from the slope of the ground have affected it, for it passed
1The Antiquity of Man, 4th edition, p. 349.
mostly over level ground or up slopes, and near its termination at Soleure the ground is actually nearly 200 feet higher than it is at the head of the lake of Geneva. There remains as a cause of motion only the slope of the upper surface of the glacier, the ice slowly flowing downward, and, by means of its tenacity and its viscocity on a large scale, dragging its lower portion still more slowly over the uneven or upward-sloping surface. This mode of motion will be discussed later when dealing with the origin of lake-basins.
No doubt at this epoch of maximum glaciation the ice-sheet extended over the whole country between the Bernese Alps and the Jura, and the downward flow of the lateral glaciers along the valley of the Sarine, Aare, and other rivers flowing towards Soleure greatly assisted the general onward motion. But the fact remains, and it cannot be too strongly insisted on, that here we have a veritable ice-sheet moving over hill and valley, carrying on its surface quantities of erratic blocks, rounding, striating, and polishing the rocks over which it passed, and with the material thus crushed and ground away forming great deposits of boulder-clay, much of which still remains, although enormous quantities must have been carried away by the rivers to the lowlands of Europe and to the sea. The fact is therefore demonstrated, and is implicitly admitted by the most conservative of glacialists, that in this case an ice-sheet has moved onward over a hilly plateau for more than 100 miles, even when its terminal moraine is at as high a level as its exit from the mountain valley where it had its origin.
Erratics in Great Britain.
It will now be well briefly to sketch the distribution of erratic blocks in Great Britain, and the conclusions to be drawn from them as to the former existence of an ice-sheet under which the greater part of our islands was buried.
Every mountain group north of the Bristol Channel was a centre from which, in the earlier and later phases of the
Ice Age, glaciers radiated; but many facts prove that during its maximum development these separate glacier systems became confluent, and formed extensive ice-sheets which overflowed into the Atlantic Ocean on the west, and spread far over the English lowlands on the east and south. This is indicated partly by the great height at which glacial striæ are found, reaching to 2,500 feet in the Lake District and in Ireland, somewhat higher in North Wales, and in Scotland to nearly 3,500 feet; but also by the extraordinary distribution of erratic blocks, many of which can be traced to localities whence they could only have been brought across the sea. The direction of the glacial striæ and of the smoothed side of ice-worn rocks also indicate that the shallow seas were all filled up by ice. The Outer Hebrides, for example, are all ice-ground from the south-east and east, showing that the deep channel of the Minch was filled up, and that the Scotch ice-sheet flowed completely over the islands. On all sides of Ireland, except the southern coast, the ice flowed outward, but on the north-east the flow was diverted southward, and on the extreme north, westward, by the pressure of the overflowing ice-sheet of Scotland which here encountered it. In like manner, the ice-marks on the east coast of Ireland and the west coast of Wales are diverted southward by the mutual pressure of their ice-sheets, which, together with that of the west of Scotland, filled up St. George's Channel. That such was the case is further proved by the fact that the Isle of Man is ice-ground in a general direction from north to south, and to the summit of its loftiest mountains which rise to a height of over 2,000 feet. This could only have been done by an ice-sheet flowing over it, and this view is further supported by some most remarkable facts in the dispersal of its local erratics. These are always found to the south of the places where they occur in situ, never to the north; and, what is still more noteworthy, they are often found far above the native rock. Thus, boulders of the peculiar Foxdale granite are found about 1,400 feet higher than
VOL. I. G
the highest point where there is an out-crop of this rock.
The Scotch ice-sheet flowed outwards on all sides, but on the east it was met by the southward extension of the great Scandinavian ice-sheet. On the extreme north the meeting of these two ice-sheets resulted in a flow to the north-west which glaciated the Orkney Islands, while the Shetlands, much farther north, received the full impact of the Scandinavian ice alone, and are therefore glaciated from the north-east. The dividing-line of the Scotch and Scandinavian ice-sheets was in the North Sea, not far from the east coast of Scotland; but farther south, at Flamborough Head and Holderness, the latter impinged on our coast, bringing with it enormous quantities of Scandinavian rocks.
Many years ago Professor Sedgwick described the cliffs of boulder-clay at Holderness as containing "an incredible number of smooth round blocks of granite, gneiss, greenstone, mica slate, &c., &c., resembling none of the rocks of England, but resembling specimens derived from various parts of the great Scandinavian chain." These are mixed, however, with a number of British rocks from the north and west, indicating the meeting ground of the two converging ice-sheets. Similar blocks occur all along the coast as far as the cliffs of Cromer in Norfolk. Across the peninsula of Flamborough about two miles west of the lighthouse there is a moraine ridge containing a few Scandinavian boulders, but mainly composed of British rocks. These latter consist of numerous carboniferous rocks from the north and north-west, together with many of Shap granite—a peculiar rock found only on Shap Fell in the eastern side of the Lake District, together with a few of Galloway granite. These facts, it will be seen, add further confirmation to the theory of great confluent ice-sheets indicated by the ice-markings upon the various groups of mountains, while it is hopelessly impossible to explain them on any theory of local glaciers, even with the aid of submergence and of floating ice.
The photo-plate here given (Fig 15) shows part of the great terminal moraine of the Scandinavian ice-sheet resting on chalk at Specton, north of Flamborough Head, 530 feet above the sea, and Mr. Kendall informs me that he has found in it characteristic Norse rocks, close to the windmill here shown. Other portions of this great moraine formed of boulder-clay, with numerous embedded ice-worn blocks of Scandinavian origin as described by Professor

FIG. 15—MORAINE AT SPECTON WITH NORSE ERRATICS.
Sedgwick, are seen in the cliffs at Thornwick, Flamborough Head, resting on chalk, (see Fig. 16 on next page), from a beautiful photograph by Mr. Bingley; and the same gentleman has given me the fine view of the cliffs of similar boulder-clay of great thickness resting on inclined strata of upper oolitic rocks further north, which, Mr. Kendall informs me, are striated from the north-east, and contain Scandinavian erratics (Fig. 17).
G 2

FIG. 16.—BOULDER-CLAY ON CHALK, FLAMBOROUGH HEAD.

FIG. 17.—BOULDER-CLAY ON UPPER OOLITE, YORKSHIRE.
Another photograph (Fig. 18) from a negative lent me by Mr. Kendall, shows a boulder of rhomb-porphyry found farther north near the mouth of the Tees, in the boulder-clay of West Rigg, 810 feet above the sea-level. It is one of the characteristic Scandinavian rocks found over a large area round the head of Christiana Fiord, Norway, but nowhere in the British Islands.

FIG. 18.—A SCANDINAVIAN BOULDER FOUND IN YORKSHIRE.
Perched Blocks.
I may here notice one of the many interesting phenomena that are only to be explained by ice-carriage, the frequent presence of what are termed perched blocks in all glaciated regions. They consist essentially of masses of rock in positions where they could not have been left by any other agency. The first of these (Fig. 19) is a large and rugged boulder of Silurian shaly rock resting upon carboniferous limestone on the Norber mountain, which forms one of the shoulders of Ingleborough. The distinctness of the erratic from the rock on which it rests is well brought out in the photo-print.
The next illustration (Fig. 20) shows a rock mass poised on the very edge of a cliff, in the pass of Llanberis, a position it could hardly have reached by any other means than ice-carriage.
Two other figures, from Falsan's La Période Glaciaire, show still more curious positions. The first (Fig. 21) is one of the great train of erratics near Monthey, Bas Valais, over 500 feet above the Rhone, standing on end upon a

FIG. 19.—SILURIAN ERRATIC ON LIMESTONE, INGLEBOROUGH.
heap of smaller blocks. The other is a mass of triassic breccia, perched on the top of two other blocks partially buried, and is perhaps the most fantastic work of the glaciers yet discovered. Its position, five miles north-west of Chambéry, is now over 160 miles from the nearest glacier (Fig. 22).
The Teaching of British Erratics.
The study of our British erratics has been assiduously pursued for many years past by a committee of the British
Association; and by means of a map showing the chief facts collected up to this date, kindly furnished me by Mr. Percy F. Kendal, secretary of the committee, I am able to give a brief sketch of the more important of the phenomena, and their bearing on the extent and motion of the British ice-sheet. The general reader may be informed that great numbers of rocks are so local and so characteristic, often being confined to a very limited

FIG. 20.—PERCHED BLOCK IN PASS OF LLANBERIS.
district or to a single mountain, that the origin of a considerable portion of the erratics can be ascertained with the greatest certainty.
Taking first the Shap granite, which has already been mentioned as occurring at Flamborough Head, we find that it has been carried northwards as far as the Solway Frith, and eastward to the Eden valley in great quantity, and over a wide area. Thence can be traced a line of boulders of this rock over the high plateau of Stainmoor into the valley of the Tees, and onward round the coast
FIG. 21.—PERCHED BLOCK, NEAR MONTHEY, BAS VALAIS.
by Scarborough to Holderness, while a branch descends southward along the valley of the Ouse to York. Coming back to its source on Shap Fell, a train of boulders of the
FIG. 22.—PERCHED BLOCK, FIVE MILES NORTH-WEST OF CHAMBÉRY.
same rock has been traced southward in a curving line, passing the east side of Morecambe Bay near Lancaster, and thence sparingly south-eastward to near Whalley. Along the same line are found boulders of peculiar granites from Eskdale and Buttermere, marking the line of junction of the northern ice-sheet with that which filled up the Irish Sea and pressed inward between the glaciers of Cumberland and North Wales. This is indicated by the fact that south of this line are scattered immense quantities of erratics, both from the south-west of Scotland and the Lake District, spreading over the whole of the low country as far as Bridgnorth and Wolverhampton, and eastward to the Derbyshire highlands. These same erratics are found round the north coasts of Wales and part of Anglesea, showing how the ice-flows divided on either side of the mountain mass of North Wales.
The centre of the great glacier sheet of North Wales appears to have been over the Arenig Mountains, whence erratics of a peculiar volcanic rock have been traced to the north and east, mingling with the last-described group; while a distinct train of these Welsh erratics stretches south-eastward to the country west of Birmingham.
In the Isle of Man are found many erratics from Galloway, and a few from the Lake District. But the most remarkable are those of a very peculiar rock found only on Ailsa Craig, a small island in the Frith of Clyde, and a single boulder of a peculiar pitchstone found only in the Isle of Arran. The Ailsa Craig rock has also been found at Moel Tryfaen on the west side of Snowdon, and more recently at Killiney, co. Dublin, on the seashore.1
The case of the boulders in the Isle of Man, which have been carried nearly 800 feet above their source, has already been mentioned; but there are many other examples of this phenomenon in our islands, and as they are of great importance in regard to the general theory of glacial motion a few of them may be noted here. So early as 1818 Mr. Weaver described a granite block on the top of Cronebane, a slate hill in Ireland, and several hundred feet higher than any place where similar granite was
1 Nature, vol. xlvii., p. 464.
to be found in situ; and he also noticed several deposits of limestone gravel in places from 300 to 400 feet higher than the beds of limestone rock which are from two to ten miles off. Débris of red sandstone is also found much higher than the parent rock. Boulders of Shap granite, Mr. Kendal tells us, have passed over Stainmoor by tens of thousands, and in doing so have been carried about 200 feet above their source; and the curious Permian rock, "Brockram," has been carried in the same direction no less than 1,000 feet higher than its highest point of origin.1 In Scandinavia there are still more striking examples, erratic blocks having been found at an elevation of 4,500 feet, which could not possibly have come from any place higher than 1,800 feet.2 We thus find clear and absolute demonstration of glacier-ice moving up-hill and dragging with it rocks from lower levels to elevations varying from 200 to 2,700 feet above their origin.
These facts have seemed so incredible that many geologists think that they throw doubt on the whole theory of a glacial epoch of the extent and nature of that indicated by the phenomena here described; while others, finding no such upward motion in any of the existing Swiss glaciers, deny that any glaciers, however large, can have produced such effects. But recent researches in Greenland and Spitzbergen have shown us exactly how such elevations of rocks and moraine-matter do actually occur. Wherever ridges or isolated rocks rise above the surface of the ice-sheet across the direction of its motion, the rock-débris that is embedded in its lower strata is gradually brought up and deposited on the surface. (See Q. Journal Geolog. Soc., Nov., 1899, p. 684, where Mr. E. J. Garwood describes this phenomenon as observed by himself in Spitzbergen.)
We see then that the ice-sheets of the British Isles, of Scandinavia, and of North America exhibit the very same characteristics as those of Switzerland, but on a larger scale; while they exhibit other phenomena which are now only to be paralleled in the great ice-sheets under which Greenland and Spitzbergen are now largely buried. We
1 Wright's Man and the Glacial Period, p. 154.
2 James Geikie's Great Ice Age, 2nd ed., p. 404.
may not be yet able to explain fully how a glacier thus moves, or what slope of the upper surface is required in order that the bottom of the ice may be forced up a given ascent, but the fact of such motion cannot any longer be denied.
The facts thus established by directed observation of existing ice-sheets, render it more easy for us to accept one of the latest conclusions of British glacialists. A great submergence of a large portion of the British Isles during the glacial period, or in the interval between successive phases of the glacial period, has long been accepted by geologists, and maps have been often published showing the small group of islands to which our country was then reduced, the supposed subsidence being about 1,400 feet. The evidence for this is the occurrence, at a few isolated spots, of glacial gravels containing marine shells in tolerable abundance, the most celebrated being at Moel Tryfaen on the west side of Snowdon, at a height of more than 1,300 feet. Shell-bearing drifts have also been found near Macclesfield at a height of over 1,100 feet, and to the east of Manchester at between 500 and 600 feet elevation. Others have since been found on Gloppa, a hill near Oswestry. The fact that the shell-bearing gravels of Moel Tryfaen are nearly 40 feet thick shows that, if they are due to submergence, the land must have remained nearly stationary at that level for a considerable period of time, and there would probably be other stationary periods at lower levels. Yet nowhere in the valleys or on the hill slopes of Wales, or the Lake District, or in the English lowlands are there any of the old beaches or sea cliffs, or marine deposits of any kind, that must have been formed during such a long continued submersion, and which can hardly have been everywhere cleared away by subsequent glaciation. Another difficulty is, that the shells of these drifts are such as could not have lived together on one spot, some being northern species, others southern, some frequenting sandy others muddy bottoms, some which live only below tidal water, while others are shore species. And, lastly, they are very fragmentary, only a small percentage of entire shells being found.
In consequence of these various difficulties it was suggested by the late Mr. Belt that the great Irish Sea ice-sheet had carried up a portion of the sea-bottom embedded in its substance, perhaps containing deposits of shells of various periods and thus explaining the inter-mixture of species as well as their fragmentary condition. The fact that boulders and pebbles from Scotland, Ailsa Craig, and Cumberland have been found in the Moel Tryfaen beds almost amounts to a proof that they were so uplifted; and a recent search has shown that in the other localities where marine shells have been found in drift at great elevations similar foreign rocks occur, rendering it almost certain that the same ice-sheets which have distributed foreign erratics so widely over our country, and which in doing so must have passed over the sea-bottom, have in a few cases carried with them a portion of that sea-bottom, and deposited it with the erratics in the places where both are now found. A full discussion of this point, with replies to various objections, by Mr. P. F. Kendal, will be found in the volume already quoted; and he has recently adduced a fresh argument against "the great submergence" in the fact that, if it ever occurred, our lowlands must for a long time have formed the bottom of a sea 200 fathoms deep, yet not a single shell characteristic of that depth has yet been discovered in the drift.1 The cumulative evidence against the submergence is now almost, if not quite, conclusive.
In the brief outline now given of the facts of glacial geology bearing upon the former existence, the thickness, extent, and motion of ice-sheets, it has only been possible to treat the subject very broadly, omitting all those details and minor difficulties which cannot be discussed within the limits of a popular exposition. My object has been to explain the nature and amount of the converging evidence demonstrating the existence of enormous ice-sheets in the northern hemisphere, to serve as a basis for the discussion of the glacial origin of certain classes of lake-basins, which will form the subject of the following chapter.
1 Wright's Man and the Glacial Period, pp. 167—175. Also Geological Magazine, November, 1892, pp 491—500.
CHAPTER V
THE ICE AGE AND ITS WORK
II. EROSION OF LAKE BASINS
LAKES are distributed very unequally over the various parts of the world, and they also differ much in their position in relation to other physical peculiarities of the surface. Most of the great continents have a considerable number of lakes, many of great size and situated on plateaux or in central basins; while the northern parts of Europe and North America are thickly strewn with lakes of various dimensions, some on the plains, others in subalpine valleys, others again high up among the mountains, these latter being of small size and usually called tarns. The three classes of lakes last mentioned occur in the greatest profusion in glaciated districts, while they are almost absent elsewhere; and it was this peculiarity of general distribution, together with the observation that all the valley lakes of Switzerland and of our own country occurred in the track of the old glaciers, and in situations where the erosive power of the ice would tend to form rock-closed basins, that appears to have led the late Sir Andrew Ramsay to formulate his theory of ice-erosion to explain them. He was further greatly influenced by the extreme difficulty or complete inadequacy of any possible alternative theory—a difficulty which we shall see remains as great now as at the time he wrote.
This question of the origin of the lake-basins of the glaciated regions is especially interesting on account of the extreme divergence of opinion that still prevails on the subject. While the general facts of glaciation, the extent
and thickness of the old glaciers and ice-sheets, and the work they did in distributing huge erratics many hundred miles from their sources and in covering thousands of square miles of country with thick layers of boulder-clay and drift, are all admitted as beyond dispute, geologists are still divided into two hostile camps when the origin of lake-basins is concerned; and the opposing forces seem to be approximately equal. Having for many years given much attention to this problem, which has had for me a kind of fascination, I am convinced that the evidence in favour of glaciation has not been set forth in all its cumulative force, while many of the arguments against it seem to me to be either illogical or beside the point at issue. I have also to adduce certain considerations which have hitherto been overlooked, but which appear to me to afford very strong, if not conclusive evidence for erosion as against any alternative theory yet proposed. I shall, therefore, first set forth, as fully as the space at my command will allow, the general evidence in favour of the ice-origin of certain classes of lakes, and the special conditions requisite for the production of lakes by this agency. The objections of the best authorities will then be considered and replied to, and the extreme difficulties of the alternative theories will be pointed out. I shall then describe certain peculiarities, hitherto unnoticed, which clearly point to erosion, as opposed to any form of subsidence or upheaval, in the formation of the lakes in question. Lastly, the special case of the Lake of Geneva will be discussed, as affording a battle-ground that will be admitted to be highly favourable to the anti-glacialists, since most of them have adduced it as being entirely beyond the powers of the ancient glaciers to have produced.
1. The different kinds of Lakes and their distribution.— To clear the ground at the outset, it may be well to state that the great plateau lakes of various parts of the world have no doubt been formed by some kind of earth movements occurring subsequent to the upheaval and partial denudation of the country. It is universally admitted that existing lakes cannot be very ancient, geologically speaking, since they would inevitably be filled up by the
sediment carried into them by the streams and by the wind. Our lakes must, therefore, be quite modern features of the earth's surface. A considerable proportion of these plateau lakes are in regions of little rainfall, and many of them have no outlet. The latter circumstance is a consequence of the former, since it indicates that evaporation either balances or exceeds the inflow. This would have favoured the formation of such lakes, since it would have prevented the overflow of the water from the slight hollow first formed and the cutting of an outlet gorge which would empty the incipient lake. Captain Dutton, in his account of the geology of the Grand Cañon district, lays stress on this fact, "that the elevation of a platform across the tract of a river rarely diverts it from its course, for the stream saws its bed into the rocks as fast as the obstacle rises." Scanty rainfall and great evaporation seem therefore to be almost essential to the formation of the larger plateau lakes. Rarely, such lakes may have been formed in comparatively well-watered districts, but the earth-movements must in these cases have been exceptionally rapid and extensive, and they are accordingly found most often in countries subject to volcanic disturbances. Such are the lakes of Southern Italy, of Macedonia, of Asia Minor, and perhaps those of Central Africa.
Quite distinct from these are the sub-alpine lakes of those mountain groups which have been subject to extreme glaciation. These are characteristically valley-lakes, occurring in the lower portions of the valleys which have been the beds of enormous glaciers, their frequency, their size, and their depth bearing some relation to the form and slope of the valleys and the intensity of the glaciation to which they have been subject. In our own country we have in Wales a small number of valley-lakes; in the Lake District, where the ice sheet can be proved to have been much thicker and to have lasted longer, we have more numerous, larger, and deeper lakes; and in Scotland, still more severely glaciated, the lakes are yet more numerous, many of those in the west opening out to the sea and forming the lochs and sounds of the Western Highlands. Coming to Switzerland which, as we have seen, bears indications of glaciation on a most
gigantic scale, we find a grand series of valley-lakes both on the north and south of the Alps, situated for the most part in the tracks of those enormous glaciers whose former existence and great development is clearly proved by the vast moraines of Northern Italy and the travelled blocks of Switzerland and France. In Scandinavia, where the ice-age reigned longest and with greatest power, lakes abound in almost all the valleys of the eastern slope, while on the west the fiords or submerged lakes are equally characteristic.
In North America, to the south of the St. Lawrence River and of Lakes Ontario and Erie, there are numbers of true valley-lakes, as there are also in Canada, besides innumerable others scattered over the open country, especially in the north, where the ice-sheet must have been thickest and have lingered longest. And in the southern hemisphere we have, in New Zealand, a reproduction of these phenomena—a grand mountain-range with existing glaciers, indications that these glaciers were recently much more extensive, a series of fine valley-lakes forming a true lake district, rivalling that of Switzerland in extent and beauty, with fiords on the south-west coast comparable with those of Norway.
Besides these valley-lakes there are two other kinds of lakes always found in strongly glaciated regions. These are alpine tarns—small lakes occurring at high elevations and very often at the heads of valleys under lofty precipices; and small or large plateau or low-level lakes which occur literally by thousands in Northern Canada, in Sweden, Finland, Lapland, and North-western Russia. The valley-lakes and the alpine tarns are admitted by all geologists to be mostly true rock-basins, though some are partially and others wholly dammed up by moraines or by land-slips; while the plateau and low-country lakes are often mere hollows in the drift with which much of the country is covered, though rock-basins are also not unfrequent.
Here, then, we see a remarkable association of lakes of various kinds with highly glaciated regions. The question is whether there is any relation of cause and effect in the association; and to determine this we must take a rapid
VOL. I. H
survey of other mountain regions where indications of ice-action are comparatively slight or altogether wanting, and see whether similar lakes occur there also. The comparison will, I think, prove very instructive.
Spain and Portugal are pre-eminently mountainous countries, there being a succession of distinct ranges and isolated mountain groups from east to west and from north to south; yet there is not a single valley-lake in the whole peninsula, but very few mountain tarns. Sardinia and Corsica are wholly mountainous, but they do not appear to possess a single valley-lake. Nor does the whole range of the Apennines, though there are many large plateau lakes in Southern Italy. Farther south we have the lofty Atlas Mountains, but giving rise to no subalpine valley-lakes. The innumerable mountains and valleys of Asia Minor have no lakes but those of the plateaux; neither has the grand range of the Lebanon, a hundred miles long, and giving rise to an abundance of rivers. Turning to the peninsula of India we have the ranges of the Ghauts, eight hundred miles long, the mountain mass of the Neilgherries and that of Ceylon, all without such lakes as we are seeking, though Ceylon has a few plateau lakes in the north. The same phenomenon meets us in South Africa and Madagascar—abundance of mountains and rivers, but no valley-lakes. In Australia, again, the whole great range of mountains from the uplands of Victoria, through New South Wales and Queensland to the peninsula of Cape York, has not a single true valley-lake. Turning now to the New World, we find no valley-lakes in the southern Alleghanies, while the grand mountains of Mexico and Central America have a few plateau lakes, but none of the class we are seeking. The extremely mountainous islands of the West Indies—Cuba, Hayti, and Jamaica—are equally deficient, although there is evidence of both upheaval and depression, in times which are geologically recent. In South America we have on the east the two great mountain systems of Guiana and Brazil, furrowed with valleys and rich in mountain streams, but none of these are adorned with lakes. And, lastly, the grand ranges of the equatorial Andes, for ten degrees on each side of the
Equator, produce only a few small lakes on the high plateaux, and a few in the great lowland river plains— probably the sites of old river channels—but no valley lakes in any way comparable with those of Switzerland or even of our own insignificant mountains.
Having thus roughly surveyed the chief mountain regions of the whole world, we find that true subalpine valley-lakes, that is, lakes in the lower parts of the valleys descending from mountain ranges or groups, filling up those valleys for a considerable distance, usually very deep, and situated in true rock-basins—that such lakes as these are absolutely unknown anywhere but in those mountain regions which independent evidence shows to have been subject to enormous and long-continued glaciation. No writer that I am acquainted with has laid sufficient stress on this really marvellous fact of lake-distribution. Professor Bonney passes it by with the remark, that there is a perfect gradation of lakes, from the smallest tarns to those of North America and Central Africa; and Mr. Douglas Freshfield says, that, wherever on the surface of our globe there are heights there must be hollows; and other writers think that lakes are general results of the process of mountain-making. But none of these writers have apparently even noticed the fact, that glacier valley-lakes have a distinctive character which separates them broadly from the lakes of all non-glaciated countries, and that they are totally absent from such countries.
But besides the mountains which possess true valley-lakes, there are a number of ranges which have been glaciated yet do not possess them, and this absence of lakes has been used as an argument against the connection of valley-lakes with glaciation. A little examination, however, shows us that these cases greatly strengthen our argument. Comparatively large and deep valley-lakes are the result of excessive glaciation, which has occurred only when conditions of latitude, altitude, and moisture combined to produce it. In regions where glaciation was of diminished intensity, from whatever causes, valley-lakes diminish in size and number, till at last only tarns are found in moderately glaciated districts. Thus, the
H 2
Pyrenees were far less severely glaciated than the Alps; they consequently possess no large valley-lakes, but numerous small high lakes and tarns. As we go eastward in the Alps, the diminished rain and snow-fall led to less severe glaciation, and we find the valley-lakes diminish in size and numbers till far east we have only tarns. The Carpathians have no valley-lakes, but many tarns. The Caucasus has no lakes, and very few tarns, and this may be partly due to the steepness of the valleys, a feature which is, as we shall see, unfavourable to lake formation. In the South Island of New Zealand the lakes are small in the north, but increase in size and number as we go south, where the glaciation was more intense. These numerous facts, derived from a survey of the chief mountains of the world, are amply sufficient to show that there must be some causal connection between glaciation and these special types of lakes. What the connection is we shall inquire later on.
2. The Conditions that favour the Production of Lakes by Ice-erosion.—Those who oppose the production of lake-basins by ice-erosion often argue as if the size of the glacier was the only factor, and urge that, because there are no lake-basins in one valley where large glaciers have been at work, those which exist in another valley where the glaciers were no larger, could not have been produced by them. But this by no means follows, because the production of a lake-basin depends on a combination of favourable conditions. In the first place it is evident that ice-erosion to some extent must have taken place along the whole length of the glacier's course, and that in many cases the result might be simply to deepen the valley all along, not quite equally, perhaps, but with no such extreme differences as to produce a lake-basin. This would especially be the case if a valley had a considerable downward slope, and was not very unequal in width or in the nature of the rocks forming its floor. The first essential to lake-erosion is, therefore, a differential action, caused locally either by increased thickness of the ice, a more open and level valley-floor, or a more easily eroded rock, or by any combination of these.
If we look at the valley-lakes of our own country and of Switzerland, the first thing that strikes us is their great length, and their situation, usually at the lower end of the valley where it emerges from the higher mountains into comparatively low country. Windermere is over ten miles long, Ullswater nearly eight miles, and the larger lakes of Switzerland and North Italy are very much longer. The first essential condition, therefore, was a valley, the lower part of which was already nearly level for several miles, and with a considerable width to the base of the mountain slopes. In the non-glaciated districts of our own country, the Dart and the Tamar are examples of rivers which have cut their valleys down nearly to sea-level while still among the hills; and in South Wales the Wye, the Usk, and the Severn have a similar character.
It must always be remembered that glacial erosion is produced by the tremendous vertical pressure of the ice, by its lower strata being thickly loaded with hard rocks frozen into its mass, and by its slow but continuous grinding motion over its bed. In the lower part of its course a glacier would be most charged with rocky débris in its under strata, since not only would it have been continually breaking off and absorbing, as it were, fresh material during every mile of its onward course, but more and more of its superficial moraines would be engulfed by crevasses or moulins, and be added to the grinding material below. That this was so is proved by the great quantity of stones and grit in the "till," which is thought by Professor James Geikie to consist, on the average, of as much stony matter as clay, sometimes one material preponderating, sometimes the other. The same thing is indicated by the enormous amount of débris often found upon the surface of the lower portions of large glaciers. The end of the great Tasman glacier in New Zealand is thus completely hidden for five miles, and most of the other glaciers descending from Mount Cook have their extremities similarly buried in débris. Dr. Diener found the Milam glacier in the Central Himalayas completely covered with moraine rubbish; and Mr. W. M. Conway states that the lowest twenty miles of the Hispar glacier (forty miles long) are "entirely covered with a mantle of moraine." If
these glaciers extended to over a hundred miles long, as did the Rhone glacier when it reached the Lake of Geneva, much of this débris would probably have found its way to the bottom, and thus supply the necessary grinding material and the abundant stones of the "till" or sub-glacial moraine-stuff, found everywhere in the tracks of the old glaciers.
Again, although ice is viscous and can slowly change its shape to almost any extent, yet it takes a considerable time to adapt itself to the continually changing outlines of a valley bottom. Hence, where great inequalities occur portions of the rocky floor might be bridged over for a considerable space, and where a valley had a narrow V-shaped bottom the sub-glacial stream might eat away so much of the ice that the glacier might rest wholly on the lateral slopes, and hardly touch the bottom at all. On a tolerably wide and level valley-bottom, however, the ice would press downwards with its fullest intensity, and its armature of densely packed stones and rock-fragments would groove and grind the rocky floor over every foot of its surface, and with a rate of motion perhaps greater than that of the existing Greenland and Alaskan glaciers, owing to the more southern latitude and therefore higher mean temperature of the soil and the ice. At the same time sub-glacial streams, sometimes forced onward under hydrostatic pressure, would insinuate themselves into every vacant groove and furrow as each graving tool successively passed on and the one behind it took a slightly different position; and thus the glacial mud, the product of the erosion, would be continually washed away, finally escaping at the lower extremity of the glacier, or in some cases getting embayed in rocky hollows where it might remain permanently as masses of clayey "till," packed with stones and compressed by the weight of the ice to the hardness of rock itself. The continual lubrication of the whole valley floor by water, together with the ever-changing form of the under surface of the glacier as it slowly moulded itself to the varying contours of the rocks beneath, would greatly facilitate the onward motion. Owing to these changes of form and the great upward pressure of the water in all the hollows to which it gained

access, it seems probable that at any one time not more than half the entire bottom surface of the glacier would be in actual contact with the rock, thus greatly reducing the total friction; while, as the process of erosion went on, the rock surfaces would become continually smoother and the inequalities less pronounced, so that even when a rock-basin had been ground out to a considerable depth the onward motion might be almost as great as at the beginning of the process.
If, now, we consider that the erosion I have attempted to describe was going on during a large part of the glacial period, under a weight of ice varying from one to five or six thousand feet in thickness, and in some cases even more; that the huge grinding tool was at work day and night, winter and summer, century after century, for whatever number of thousands of years we give to the glacial period; that—as innumerable other facts prove—the ice moved irresistibly over hill and dale, and up slopes far steeper than any formed by the inclines of the bottoms of our deepest lakes— as shown by the true scale section of Lake Como here given — what is there of impossible, or even of improbable in the belief, that lake-basins were produced by such differential erosion? To the ordinary observer it seems impossible that a mountain valley, half a mile wide and bounded by rocky slopes and precipices two or three thousand feet high, can have been formed without any "convulsion of nature," but merely by the natural agencies he sees still in action—rain and frost, sun and wind—and that the small rock-encumbered stream now flowing along its bottom can have
carried away the whole of the many cubic miles of solid rock that once filled up the valley. But the geologist knows that these apparently insignificant forces have done the work, through their continuous action always in one direction for thousands, or even for millions of years; and, therefore, having before him so many proofs of the eroding power of ice, in planed and rounded rocks and in the grooves and furrows which are the latest marks left by the ice-tool; and bearing in mind the long duration and possibly recurrent phases of the ice age— to be measured certainly by tens, perhaps by hundreds, of thousands of years—he can have little difficulty in accepting the erosion of lake-basins as the most satisfactory explanation of their origin.
3. Objections of Modern Writers considered.—Professor Bonney and many other writers ask, why lakes are so few though all the chief valleys of the Alps were filled with ice; and why, for instance, there is no great lake in the Dora Baltea valley whose glacier produced the great moraines of Ivrea opposite its outlet into the plains of Italy, and which form a chain of hills fifteen miles long and fifteen hundred feet high? The answer, in the case of the Dora Baltea is not difficult, since it almost certainly has had a series of lake-basins at Aosta, Verrex, and other places where the broad level valley is now filled with alluvial gravel. But the more important point is the extreme narrowness of the lower part of the valley above Donnas and again near its entrance into the valley of the Po. The effect of this would be that the great glacier, probably two thousand feet thick or more, would move rapidly in its upper layers, carrying out its load of stones and débris to form the terminal moraine, while the lower strata, choked in the defiles, would move very slowly. And once out in the open valley of the Po, then a great inlet of the warm Mediterranean Sea, the ice would rapidly melt away in the water and in the warm moist atmosphere, and therefore have no tendency to erode a lake-basin.
The Lake of Lugano, with its curious radiating arms, is said to be another difficulty. But each of these arms is the outlet of a valley or series of valleys, which were
no doubt reduced to nearly level plains by sub-aerial denudation before the ice began its work. The basin of these valleys comprises about two hundred square miles and the watershed to the north is moderately high: but there can be no doubt that a large overflow from the Como glacier poured into it; and the difficulty seems to me to be purely imaginary if we simply recognise the fact that an essential preliminary to lake-erosion is a pre-existing nearly level valley bottom.
Another difficulty is said to be the frequent presence of islands in the lakes; but here again the answer is easy. The islands, always ground down to roches moutonnées, were craggy hills in the pre-existing valleys, and such hills existed because they had for ages resisted the sub-aerial denudation which had hollowed out the valleys. The same characters of density or toughness that enabled them to resist ordinary denudation, enabled them also, to some extent, to resist destruction by ice-erosion; just as the character of the rocks which enabled ordinary denudation to bring them down to a nearly level surface in the valley-bottom, also facilitated the ice-erosion which converted the level valley floor into a rock-basin, and after the ice left it into a lake.
Every writer brings forward the well-known fact that the ends of glaciers pass over beds of gravel or moraine matter, without destroying or even disturbing it. But there is no reason why they should do more than compress such beds of loose material and roughly level their surfaces. It is the old delusion of a glacier acting like a scoop or a plough that leads to the idea that if it can erode rock slowly it must altogether demolish gravel or boulder-clay. But if we turn to the description I have given of how a glacier erodes a rock-basin and apply this to its passage over a bed of gravel or boulder-clay, we shall see that in the latter case the erosion would be much more difficult, because each ice-embedded stone or rock would press into the yielding material, which would close up instantly behind it under pressure of the ice and thus leave no result. Where the sub-glacial water accumulated, channels would be cut in the gravel or clay, but elsewhere there would
probably be no erosion at all. Some writers maintain that the lakes were all filled up with alluvium previous to the glacial epoch, and that the ice cleared out this incoherent matter; but it is almost certain that no such clearance would have taken place, because the glacier would pass over such a surface, the stones temporarily furrowing it, while the sub-glacial water would cut for itself one or more deep channels, and there would thus be no water flowing over the whole surface of the basin, which must be so great an aid to erosion in solid rock.
These considerations apply to the equally common objection, that the great masses of boulder-clay left behind by the ice-sheet, and over which it must have passed, prove that it could have had little eroding power. The product of the erosion of irregular rock-surfaces in an undulating tract of country, where not carried away by water, would necessarily, by the pressure of the ice, be forced into the more or less sheltered or land-locked hollows, thus tending to equalise the surface-contours and facilitate the onward motion of the ice. In such hollows it would be pressed and compacted by the weight of the ice, but would be neither eroded nor forced away until, by the continued process of rock-erosion, it became exposed to unequal lateral pressure, when it would be gradually removed to some other sheltered hollow, perhaps to again undergo the same process of removal at a later period, and finally rest in the positions in which we find it. During the later stages of the ice age when, notwithstanding the onward motion of the middle portions of the glacier, the lower portion was melting away both above and below, and the terminal ice-cliff was permanently retreating, almost the whole of the eroded matter except what was carried away by the sub-glacial torrents, would remain behind; and it is this final product of glacial erosion that forms the huge deposits of boulder-clay which encumber the surface of the lowlands in most highly glaciated countries. When, however, the moving ice changed its direction, as it often did during the varying phases of the ice age, it sometimes acted most energetically in crushing, dragging, and contorting both the boulder-clay and other
superficial beds, often causing the wildest confusion in the deposits and sometimes embedding huge sheets of tertiary strata or chalk in the midst of the boulder-clay. But this is a very different mode of action from that by which hard rocks are ground down or lake-basins eroded.
In reply to the continual assertions of Professor Bonney and of most of the Alpine explorers, that the action of glaciers is entirely superficial, and that they actually preserve the surfaces they cover from denudation, a few facts may be here given. From a large number of gaugings by Dollfus-Ausset, Dr. Penck has calculated that the solid matter in the torrent which issues from the Aar glacier annually amounts to six hundred and thirty-eight cubic metres for each square kilometre of the surface of the glacier, a quantity sufficient to lower the bed of the glacier one metre in sixteen hundred and sixty-six years, or one foot in five hundred and five years; and the same writers calculate that the same amount of erosion in a valley by water alone would require two and a half times as long.1 Other writers have made estimates less favourable to ice as an agent of erosion; but even if the amount annually ground away be but small, the cumulative effect was undoubtedly very great in the case of the enormous glaciers of the ice age. The very wide areas covered with boulder clay and drift in North America, and its great average depth, have already been referred to in the preceding chapter (p. 75); but a still more striking estimate has been made of the amount of rock débris in Northern Europe which can be traced to Scandinavia. Dr. Amund Helland states that about eight hundred thousand square miles are covered with such drift to an average depth of one hundred and fifty feet, of which about one hundred feet are of Scandinavian origin, the remainder being local. The area of Scandinavia and Finland, from which this débris has been derived, is very much less than the area over which it is distributed, so that to produce it an amount equal to an average thickness of two hundred and fifty-five feet must have been removed from those countries. To this must be added the amount which has gone
1 Falsan, La Période Glaciaire, p. 90.
into the Baltic and North Seas, and also that which has been carried away by rain and rivers since the ice age passed away, and yet further, the enormous amount that still remains on the lowlands of Scandinavia, and we shall then arrive at an amount probably twice as great as the above estimate, that is, something like five hundred feet as the average amount of denudation of Scandinavia during the glacial period.1 Now, unless this estimate is wildly and extravagantly erroneous—and Professor Geikie adopts it as, primâ facie, not extravagant, we have an amount of ice-erosion so enormous as to put completely out of court all the allegations of those who attempt to minimise it as a mere smoothing off of sharp angles and rugged surfaces. I am not aware that Professor Bonney denies the Scandinavian origin of the greater part of the northern drift, and unless he can show that its quantity is something like a fiftieth part only of the estimate of Dr. Helland, I cannot understand how he can still maintain that the glaciers and ice-sheets of the ice age were agents of abrasion, not of erosion, and that they were therefore impotent to grind away the comparatively small amount of rock removed, under the most favourable conditions, from the basins of the valley-lakes whose origin we are discussing.
Of course, it may be said that the bulk of the Scandinavian drift in Northern Europe is not a result of ice-erosion, but of sub-aerial denudation, the product of which was carried away by the ice-sheet. This may be admitted as regards some portion of it, and especially of the heaps of angular blocks previously referred to. There was also probably a considerable layer of soil and of partially disintegrated rock in Scandinavia before the glacial epoch, but both these elements will certainly not account for more than a small portion of the drift, which consists largely of boulder-clays, the undoubted product of glacial erosion. This question has been carefully examined by glacialists, and Professor James Geikie's conclusion is as follows, referring specially to this northern drift:—
"It is obviously impossible that the ground-moraines of an ice-sheet of such dimensions could have been derived or even supple-
1 Fragments of Earth Lore, by James Geikie, F.R.S. (1893), p. 167.
mented to any extent from superficial sources. The boulder clays are the direct products of glacial erosion. They consist essentially of unweathered material. Boulders, smaller stones, grit, sand, and the finer grained rock-meal or flour are all alike fresh, they have not been altered chemically as they would have been had they come from superficial sources. They could not have come from above, and they cannot represent the weathered rock-débris of preglacial times." (Earth Sculpture, p. 187.)
And again (at p. 190 of the same volume) he writes:—
"From the various facts above referred to we are justified in concluding that glacier-ice is a most effective agent of erosion. It not only abrades, rubs, smooths, and polishes, but crushes, folds, disrupts, and displaces rock-masses, the amount of disturbance being in proportion to the resisting power of the rock and the pressure exerted by the ice. Other things being equal, more crushing and displacement will be effected under a massive ice-sheet than under a small valley glacier."
Taking these various facts and arguments in connection with the great number and large size of the lakes of Sweden, Lapland, and Finland, almost all true rock basins, the cumulative evidence in favour of the origin of such lakes by glacial erosion becomes overwhelming.
4. The Alternative Theory and its Difficulties.—There is really only one alternative theory to that of ice-erosion for the origin of the class of lakes we have been discussing, viz., that they were formed a little before the glacial epoch or during its continuance, by earth-movements of the same nature as those which are concerned in mountain formation, that is, by lateral pressure causing folds or flexures of the surface; and where such flexures occurred across a valley a lake would be the result. This is Professor Bonney's theory given in his paper in the Geographical Journal, and it is also that of Desor, Forel, Favre, and other eminent continental geologists. It is explained fully in the work of M. Falsan (already quoted), who also adopts it; and it may be considered, therefore, that if this theory can be shown to be untenable that of glacial erosion will hold the field, since there is no other that can seriously compete with it. Professor Bonney considers this theory completely satisfactory, and he complains that the advocates of glacial
erosion have never discussed it, intimating that they "deemed silence on this topic more prudent than speech."
As this theory is put forward with so much confidence, and by geologists of such high reputation, I feel bound to devote some space to its consideration, and shall, I think, be able to show that it breaks down on close examination.
In the first place, it does not attempt to explain that wonderful absence of valley-lakes from all the mountain regions of the world except those which have been highly glaciated. It is, no doubt, true that during the time the lakes were filled with ice instead of water they would be preserved from filling up by the influx of sediment; and this may be fairly claimed as a reason why lakes of this class should be somewhat more numerous in glaciated regions, but it does not in any way explain their total absence elsewhere. We are asked to believe that from the period immediately preceding the glacial epoch to our own day, earth-movements of a nature to produce deep lakes occurred in every mountain range without exception that was about to be or had been subject to severe glaciation, and not only so, but occurred on both sides of each range, as in the Alps, or all round a mountain range, asin our Lake district, or in every part of a complex mountain region, as in Scotland from the Frith of Clyde to the extreme north coast—all in this very limited period of geological time. We are further asked to believe that during the whole period from the commencement of the ice-age to our day such earth-movements have never produced a single group of valley-lakes in anyone of the countless mountain ranges and hilly regions throughout the whole of the very much more extensive non-glaciated regions of the globe! This appears to me to be simply incredible. The only way to get over the difficulty is to suppose that earth-movements of this nature occurred only at that one period, just before the ice age came on, and that the lakes produced by them in all other regions have since been filled up. But is there any evidence of this? And is it probable that all lakes so produced in non-glaciated regions, however large and deep they might be, and however little sediment was carried down by their inflowing streams, should have entirely
disappeared? The theory of the pre-glacial or inter-glacial origin of these lakes by earth movements thus rests upon a series of highly improbable suppositions entirely unsupported by any appeal to observed facts.
There is, however, another difficulty which is perhaps even greater than those just considered. Whatever may be the causes of the compression, elevation, folding, and other earth-movements which have led to the formation of mountain masses, there can be no doubt that they have operated with extreme slowness; and all the evidence we have of surface movements now going on show that they are so slow as to be detected only by careful and long-continued observations. On the other hand, the action of rivers in cutting down rocky barriers is comparatively rapid, especially when, as in all mountainous countries, they carry in their waters large quantities of sediment, and during floods bring down also abundance of sand, gravel, and large stones. A remarkable illustration of this erosive power is afforded by the river Simeto in Sicily, which has cut a channel through solid lava which was formed by an eruption in the year 1603. In 1828, Sir Charles Lyell states, it had cut a ravine through this compact blue rock from fifty to several hundred feet wide, and in some parts from forty to fifty feet deep.1 The enormous cañon of the Colorado, from 3,000 to 5,000 feet deep and 400 miles long, which has been entirely cut through a series of mesozoic and palæozoic rocks during the latter portion of the tertiary period, is another example of the wonderful cutting power of sand-laden running water.
It is, in fact, only on account of this powerful agency that we do not find valley lakes abounding in every mountainous country, since it is quite certain that earth-movements of various kinds must have been continually taking place. But if rivers have always been able to keep their channels clear, during such movements, among the mountains of the tropics and of all warm countries, some reason must be found for their inability to do so in the Alps and in Scotland, in Cumberland, Wales, and southern New Zealand; and as no reason is alleged, or any proof offered
1 Principles of Geology, 11th ed., vol. i., p. 353.
that sufficiently rapid and extensive earth-movements actually did occur in the subalpine valleys of these countries at the epoch required we must decline to accept such a hypothetical and unsatisfactory explanation.
Nothing is more easy, and nothing seems at first sight more plausible, than to allege these "earth-movements" to account for any one lake whose origin may be under discussion. But it ceases to be either easy or plausible when we consider the great number of the lakes to be accounted for, their remarkable positions and groupings, and their great depths. We must postulate these movements, all about the same time, in every part of the Highlands of Scotland, everywhere in the Lake district, and on both sides of the Alps. Then, again, the movements must have been of greater extent just where we can prove the glaciation to have been most severe. It produced lakes from 100 feet to 270 feet deep in Cumberland and Westmoreland; in Scotland, where the ice was much thicker, the lakes are from over 300 to over 1,000 feet deep; while in the Alps of Switzerland and North Italy, with its vast glaciers and ice-sheets, many are over 1,000 feet, and one reaches the enormous depth of over 2,500 feet. It may be said that the depth is in proportion to the height of the mountains; but in equally high mountains that have not been glaciated there are no lakes, so this cannot be the true explanation. One more remarkable coincidence must, however, be pointed out. The two largest Swiss lakes—those of Geneva and Constance—are situated just where the two greatest West European rivers, the Rhone and the Rhine, get beyond the mountain ranges; while on the south, one of the largest and by far the deepest of the lakes—Lago Maggiore—collected into its basin the glacier streams from a hundred miles of the high Alps, extending from Monte Rosa on the west to the peaks above San Bernardino on the east. Throughout this great curve of snowy peaks the streams converge, with an average length of only 30 miles, to unite in a valley only 646 feet above the sea level. No such remarkable concentration of valleys is to be found anywhere else in the Alps, and no other lake reaches to nearly so great a depth.
On the theory of glacial erosion we have here cause and effect; on that of earth movements we have another mere coincidence added to the long series already noticed. The depth of over 2,500 feet undoubtedly seems enormous, but that depth exists just at the point where the two great valleys which have collected the converging streams above referred to, unite together. Geologists will probably not think thirty thousand years an extravagant estimate for the duration of the glacial period, in which case an erosion of only an inch in a year would be sufficient. Lago di Garda, the largest Italian lake, had a still larger catchment area in glacial times, but not nearly so much concentrated, hence, perhaps, its comparatively moderate depth of about 1,000 feet. We see, then, that on the theory of erosion, the size, depth, and position of the chief lakes are all intelligible, while on that of earth movements they have no meaning whatever, since the deep-seated agencies producing subsidence, upheaval, or curvature of the surface would be as likely to act in the small as in the large valleys, and to produce deep lakes in other places than those where, at a later epoch, the thickest glaciers accumulated.
5. The Contours and Outlines of the Lakes indicate Erosion rather than Submergence.—While collecting facts for the present articles, it occurred to me that the rival theories of lake-formation—erosion and submergence—were so different in their mode of action that they ought to produce some marked difference in the result. There must be some criteria by which to distinguish the two modes of origin. Under any system of earth movements a valley-bottom will simply become submerged, and be hardly more altered than if it had been converted into a lake by building an artificial dam in a convenient situation. We should find, therefore, merely a submerged valley with all its usual peculiarities. If, however, the lake-basin has been formed by glacial erosion, then some of the special valley-features will have been destroyed, and we shall have a distinct set of characters which will be tolerably constant in all lakes so formed. Now I find that there are three such criteria by which we ought to be able to dis-
VOL. I. I
tinguish the two classes of lakes, and the application of these tests serves to show that most of the valley-lakes of glaciated countries were not formed by submergence.
The first point is that valleys in mountainous countries often have the river-channel forming a ravine for a few miles, afterwards opening out into a flat valley, and then again closing, while at an elevation of a hundred or a few hundred feet, at the level of the top of the ravine, the valley walls slope back on each side, perhaps to be again flanked by precipices. Now, if such a valley were converted into a deep lake by any form of subsidence, these ravines would remain under water and form submerged river channels. But neither in the lakes which have been surveyed by the Swiss Government, nor in the Atlas des Lacs Françaises of M. Delebecque, nor in those of the German Alps by Dr. Alois Geistbeck, nor in the lakes of our own country, can I find any indications of such submerged river-channels or ravines, or any other of the varied rock features that so often occur in valleys. Almost all these lakes present rather steeply sloping sides with broad, rounded, or nearly level bottoms of saucer shape, such as are certainly not characteristic of sub-aerial valley bottoms, but which are exactly what we might expect as the ultimate result of thousands of years of incessant ice-grinding. The point is, not that the lake-bottoms may not in a few cases represent the contours of a valley, but that they never present peculiarities of contour which are not unfrequent in mountain valleys, and never show submerged ravines or jutting rocky promontories like those which are so common a feature in hilly districts.
The next point is, that Alpine lake-bottoms, whether large or small, frequently consist of two or more distinct basins, a feature which could not occur in lakes due to submergence unless there were two or more points of flexure for each depression, a thing highly improbable even in the larger lakes and almost impossible in the smaller. Flexures of almost any degree of curvature are no doubt found in the rocks forming mountain chains; but these flexures have been produced deep down under enormous pressure of overlying strata, whereas the surface beds
which are supposed to have been moved to cause lakes are free to take any upward or downward curves, and as the source of motion is certainly deep-seated those curves will usually be of very gradual curvature. Yet in the small lake of Annecy there are two separate basins; in Lake Bourget also two; in the small lake of Aiguebellette, in Savoy, there are three distinct basins of very different depths; and in the Lac de St. Point, about four miles long, there are also three separate flat basins. In Switzerland the same phenomenon is often found. In the Lake of Neufchatel there are three basins separated by ridges from twenty to thirty feet above the deeper parts. The small Lac de Joux, at the head of a high valley in the Jura, has also three shallow basins. Lake Zurich consists of three well-marked basins. The exceedingly irregular Lake of Lucerne, formed by the confluence of many valleys meeting at various angles hemmed in by precipitous mountains, has eight distinct basins, mostly separated by shallows at the narrow openings between opposing mountain ridges. This is exactly what would result from glacier action, the grinding power of which must always be at a maximum in the wider parts of valleys, where the weight of the ice could exert its full force and the motion be least impeded. On the subsidence or curvature theory, however, there is no reason why the greatest depth should occur in one part rather than in another, while separate basins in the variously diverging arms of one lake seem most improbable. The lakes of Thun and Brienz form two basins of what was evidently once a single lake. The upper or Brienz basin is enormously deep, over two thousand feet, and the reason is obvious. The combined glaciers of the Lauterbrunnen and Grindelwald valleys entered the main valley in a direction almost opposite to that of the Aare, piling up the ice against the great barrier of the Rieder Grat, so that it at length flowed downward with greatly increased grinding power; while lower down, towards Thun, the valley opens widely and would thus allow the ice to spread out with greatly diminished thickness. In our own country Loch Lomond and Ullswater have been found to consist of several distinct basins, and in none
I 2
of our lakes have any indications of submerged river channels yet been found.
The third point of difference between lakes of erosion and those of submersion is the most important and the most distinctive, and furnishes, I think, what may be termed a diagnostic character of lakes of erosion. In most river valleys through a hilly or mountainous country outside of the glaciated districts, the tributary streams entering more or less at right angles to the main valley are seen to occupy small valleys of their own, which usually open out for a short distance at the same level before joining the main valley. Of course there are also torrents which rush down steep mountain slopes directly to the main river, but even these have usually cut ravines more or less deeply into the rock. Now if in such a valley we could mark out a contour line two hundred, three hundred, or five hundred feet above the level of the main stream, we should see that line continually turning up each side valley or ravine till it reached the given level at which to cross the tributary stream, and then turning back to the main valley. The contour line would thus form a series of notches or loops of greater or less depth at every tributary stream with its entering valley or deeply cut ravine, and if the main valley were filled with water this line would mark out the margin of the lake. As an illustration of this feature we may take the south-west coast of England, which has never been glaciated, but which has undergone a slight recent subsidence as indicated by the submerged forests which occur at several places. The result of this submergence is that the lower parts of its larger river valleys have been converted into inland tidal lakes, such as Poole Harbour, Dartmouth Harbour, Kingsbridge River, Plymouth and Devonport Harbours, and Carrick Road above Falmouth. The Dart River is an excellent example of such a submerged valley, and its outline at high-water mark is shown at (3) on the accompanying cut, where the characteristic outline of such a valley is well indicated, the water running up every tributary stream as described above. The lower section (4) shows the same feature by means of a map of the River
Lake Forms due to Erosion (1, 2), to Submersion (3, 4).
Tweed, near Peebles, with the seven hundred feet contour line marked on it by a dotted line.1 If the valley were submerged to this depth the dotted line would mark the outline of a lake, with arms running up every tributary stream just as in the case of the river Dart. Although situated in a glaciated district the valley here is postglacial, all the old river channels being deeply buried in drift.
If we now turn to the valley-lakes in glaciated districts we shall find that they have a very different contour, as shown by the two upper outline maps on the same page; (1) showing the upper part of Ullswater on a scale of one mile to an inch, as in the Dart and Tweed maps, and (2) showing the upper part of Lake Como, taken from the Alpine Club map, on a scale of four miles to an inch. In both of these it will be seen that the water never forms inlets up the inflowing streams, but all of these without exception form an even junction with the lake margin, just as they would do if flowing into a river. Exactly the same feature is present in the lower portions of these two lakes, and it is equally a characteristic of every lake in the Lake district, and of all the Swiss and Italian lakes. On looking at the maps of any of these lakes one cannot but see that the lake surface, not the lake bottom, represents approximately the level of the pre-glacial valley, and that the lateral streams and torrents enter the lake in the way they do because they could only erode their channels down to the level of the old valley before the ice overwhelmed it. Of course this rule does not apply to large tributary valleys carrying separate glaciers, since these would be eroded by the ice almost as deeply as the main valley.
The three features of the valley-lakes of glaciated regions now pointed out—the absence of submerged ravines or river-channels either of the main river or of tributary streams; the basin-forms of the lake bottoms and the frequent occurrence of two or more separate basins even in small lakes; and the simple form of surface con-
1 Copied from a portion of the map at p. 144 of Geikie's Great Ice Age, taken from the Ordnance Survey Map.
tour of all this class of lakes, so strongly contrasting with that of valleys known to have been recently submerged, as well as with the contour lines of valleys in non-glaciated districts and in those which are known to be post-glacial—seem to afford, as nearly as the case admits, a demonstration that the lakes presenting these features have been formed by erosion and not by submergence.
It may, of course, be urged that the reason the lakes in question exhibit smooth contours is, because the lateral inlets which once existed have become filled up with sediment. In some, perhaps in many, cases this may be the explanation, but it can hardly be so in all, considering the very limited amount of material brought down by many of the smaller rocky streams, and the great depth to be filled up in the case of the deeper lakes. In Lake Como, for example, a few of the lateral streams have formed small projecting deltas, but they evidently run in steep rock beds to the contour edge of the lake, not one of them showing any sign of having had a submerged ravine now filled up with alluvium. And the same is the case with the lakes of Geneva, Brienz, and most others. When lateral alluvial valleys at all commensurable in size with the depth of the lake are proved to exist, this objection may be made. At present it is a mere supposition.
In connection with this subject may be noticed the many cases in which Alpine valleys present indications of having been greatly deepened by glacial erosion, although, owing either to the slope of the ground or the uniformity of the ice-action, no lake has been produced. In some valleys, as in that of Lauterbrunnen, the trough between the vertical rock-walls was probably partly formed before the ice age, but was greatly deepened by glacial erosion, the result being that the tributary streams have not since had time to excavate ravines of equal depth with the main valley, and therefore form a series of cascades over the lateral precipices, of which the Staubbach is the finest example. In many other cases, however, the side streams have cut wonderfully narrow gorges by which they enter the main valley. This work was probably begun by a sub-glacial stream, and the action of the
atmosphere being shut out by the superincumbent ice and all variation of temperature avoided, the torrent cut for itself a very narrow groove, sometimes with overhanging sides, as it found layers of somewhat softer rock to eat away; and the upper surface of the rock being ground smooth by the ice, the atmosphere has had little effect since, and the gorge, while deepened below, has remained as restricted above as when first eroded. Such are the gorges of the Trient, Leuk, Pfäffers, and many others well known to Alpine tourists. I am not aware whether such extremely narrow winding gorges, often only two or three feet between the rock walls, are to be found in countries which have never been glaciated. I do not myself remember reading of any, though of course tremendously deep ravines are common, but these are of quite a different character. Should it be found that these extremely narrow rock-walled gorges are peculiar to glaciated districts they will afford us a means of estimating the amount of glacial erosion in valleys where no lake basins have been formed.1
The Lake of Geneva as a Test of the Rival Theories.—When I recently began to study this question anew, I was inclined to think that the largest and deepest of the Alpine lakes, such as Geneva, Constance, Lago Maggiore, and Lago di Garda, might perhaps have originated from a combination of earth-movements with ice-erosion. But on further consideration it appears that all the characteristic features of erosion are present in these as fully as in the smaller lakes. They are situated in the largest river-valleys or in positions of greatest concentration of the glacier-stream; their contours and outlines are those of eroded basins; while all the difficulties in the way of an origin by earth-movements are as prominent in their case as in that of any other of the lakes. I will therefore discuss, first, some of the chief objections to the erosion theory as applied to the above named lake, and then consider the only alternative theory that has obtained the acceptance of modern writers.
One of the first objections made was, that the lake did not lie in the direction of the greatest action of the glacier,
1 Some evidence on this point will be found in the next chapter.
which was straight across to the Jura where the highest erratic blocks are found. This was urged by Sir Charles Lyell immediately after Ramsay's paper was read, and as it has quite recently been put forth by Professor Bonney, it would appear to be thought to be a real difficulty. Yet a little consideration will show that it has not the slightest weight. The lake was not eroded in the line of motion of the central and highest part of the old glacier, because that line was over an elevated and hilly plateau, which is even now from five hundred to a thousand feet above the lake, and was then even higher, since, the ice-sheet certainly effected some erosion.1 The greatest amount of erosion was of course in the broad and nearly level valley of the pre-glacial Rhone, which followed the great curve of the existing lake, and had produced so open a valley because the rocks in that direction were easily denuded. Objectors invariably forget or overlook the indisputable fact that the existence of a broad, open, flat-bottomed valley in any part of a river's course, proves that the rocks were there either softer or more friable, or more soluble, or by some combination of characters more easily denuded. A number of favourable conditions were combined to render ice-erosion easy in such a valley. The rock was, as we have shown, more easy to erode; owing to the low level the ice was thicker and had greater weight there than elsewhere; owing to the flatness and openness of the valley the ice moved more freely there; owing to the long previous course of the glacier its under surface would be heavily loaded with rock and grit, which, during its whole course would, by mere gravitation, have been slowly working its way downward to the lowest level; and, lastly, all the sub-glacial torrents would accumulate in this lowest valley, and, as erosion went on, would, perhaps under hydrostatic pressure, wash away all the ground-out material and so facilitate erosion. To ask why the lake was formed in the valley, where everything favoured erosion, rather than on the plateau where everything was against it, is to make mere verbal objections which have no relation to the conditions that actually existed.
1 See Map at p. 128.
Another objection almost equally beside the real question is to ask why the deepest part of the lake is near the south or convex side, whereas a stream of water always exerts most erosive force against the concave side.1 The answer is, that ice is not water, and that it moves so slowly as to act, in many respects, in quite a different manner. Its greatest action is where it is deepest—in the middle of the ice stream—while water acts least where it is deepest, and more forcibly at the side than in the middle. The lake is, no doubt, deepest in the line of the old river, where the valley was lowest; and that may well have been nearer the southern than the northern side of the lake.
Another frequently urged objection is, that as the glacier has not widened the narrow valley from Martigny to Bex it could not have eroded a lake nearly a thousand feet deep. This seems to me a complete non sequitur. As a glacier erodes mainly by its vertical pressure and by the completeness of its grinding armature of rock, it is clear that its grinding power laterally must have been very much less than vertically, both on account of the smaller pressure because it would mould itself less closely to the ever-varying rocky protuberances, and mainly, perhaps, because at the almost vertical sides of the valley it would have a very small stony armature, the blocks continually working their way downward to the bottom. Thus, much of the ice in contact with the sides of narrow ravines might be free of stones, and would therefore exert hardly any grinding power. It is also quite certain that the ice in this narrow valley rose to an enormous height, and that the chief motion and also the chief erosion would be on the lateral slopes, while the lower strata, wedged in the gorge, would be almost stationary.
The most recent researches, according to M. Falsan, show that the thickness of the ice has been usually underestimated. A terminal moraine on the Jura at Chasseron is 4,000 feet above the sea, or 2,770 feet above Geneva. In order that the upper surface of the ice should have had
1 Falsan, La Période Glaciaire, p. 153. Fabre, Origine des Lacs Alpins, p. 4.
sufficient incline to flow onward as it did, it was probably 5,000 or 6,000 feet below Martigny and 4,000 or 5,000 feet over the middle of the lake. It is certain, at all events, that whatever thickness was necessary to cause onward motion that thickness could not fail to be produced, since it is only by the onward motion to some outlet or lowland where the ice can be melted away as fast as it is renewed that indefinite enlargement of a glacier is avoided. The essential condition for the formation of a glacier at all is, that more ice should be produced annually than is melted away. So long as the quantity produced is on the average more than that melted, the glaciers will increase; and as the more extended surface of ice, up to a certain point, by forming a refrigerator helps its own extension, a very small permanent annual surplus may lead to an enormous extension of the ice. Hence, if at any stage in its development the end of a glacier remains stationary, either owing to some obstacle in its path or to its having reached a level plain where it is unable to move onward, the annual surplus of ice produced will go to increase the thickness of the glacier and its upper slope till motion is produced. The ice then flows onward till it reaches a district warm enough to bring about an equilibrium between growth and dissolution. If, therefore, at any stage in the growth of a glacier a thickness of six, seven, or even eight thousand feet is needed to bring about this result, that thickness will inevitably be produced. We know that the glacier of the Rhone did move onward to the Jura and beyond it; that the northward branch flowed on beyond Soleure till it joined the glacier of the Rhine; and that its southern branch carried Alpine erratics to the country between Bourg and Lyons, more than 200 miles from its source. We know, too, that throughout this distance it moved at the bottom as well as at the top, by the rounded and polished rocks and beds of stiff boulder clay which are found in almost every part of its course.
In view, therefore, of the admitted facts, all the objections alleged by the best authorities are entirely wanting in real force or validity; while the enormous size
and weight of the glacier and its long duration, as indicated by the great distance to which it extended beyond the site of the lake, render the excavation by it of such a basin as easy to conceive as the grinding out of a small alpine tarn by ice not one-fourth as thick, and in a situation where the grinding material in its lower strata would probably be comparatively scanty.
We have now to consider the theory of Desor, adopted by M. Favre, and set forth in the recent work of M. Falsan as being "more precise and more acceptable" than that of Ramsay. We are first made acquainted with a fact which I have not yet alluded to, and which most writers on the subject either fail to notice or attempt to explain by theories, as compared with which that of Ramsay is simple, probable, and easy of comprehension. This fact is, that around Geneva at the outlet of the lake, as well as at the outlets of the other great lakes, there is spread out an old alluvium which is always found underneath the boulder-clay and other glacial deposits. This alluvium is, moreover, admitted to be formed in every case of materials largely derived from the great Alpine range. Now here is a fact which of itself amounts to a demonstration that the lakes did not exist before the ice age; because, in that case all the Alpine débris would be intercepted by the lake (as it is now intercepted) and the alluvium below the glacial deposits would be, in the case of Geneva, that formed by the wash from the adjacent slopes of the Jura; while in every case it would be local not Alpine alluvium.
Professor James Geikie informs me that he considers the so-called "old alluvium" to be probably only the fluvio-glacial gravels and sands swept out from underneath the advancing glacier, and therefore to be no older, geologically, than the moraine matter which overlies it. The Swiss geologists, however, do not appear to hold this view, since they have recourse to a very remarkable hypothesis in order to overcome what they evidently believe to be a real difficulty in the way of the preglacial origin of the lake. The suggested explanation is as follows: At the beginning of the ice age the glacier
of the Rhone crept on down its valley past Martigny and St. Maurice till it reached the lake; it is then supposed not to have marched on with an ice-wall, say five hundred or more feet high, but to have at once spread out like so much soft pitch, and to have filled the lake to its present water-level or thereabouts. Then, over this great plain of ice, the sub-glacial torrent of the Rhone is supposed to have flowed, carrying with it and depositing at the end of the lake that ancient alluvium which, somehow, has got to be accounted for!1
Having thus filled the lake with ice instead of water, the main body of the glacier is supposed to start afresh and to travel over the ice, and thus obviate the imaginary difficulty of a glacier moving up hill, though every student of glaciers now admits that they did so, and though it is universally acknowledged that this very glacier of the Rhone moved over higher, steeper, and more irregular hills on its way to the Jura and to Soleure.
Now this extraordinary theory involves two difficulties which are passed by in silence, but which seem to entirely contravene all that we know of the nature of glaciers, and to be entirely unsupported by facts. The first is—the glacier ceasing to move onward as a glacier, but spreading out to fill up a lake basin, as if the lake were simply frozen to the bottom. Is this conceivable or possible? I think not. When glaciers come down to a fiord or to the sea they do not spread out laterally, but move on till the water is deep enough to buoy them up and break off icebergs, and no reason is given why anything different should have happened in the case of the great Swiss and Italian lakes, supposing they existed before the ice age came on. That the glacier should afterwards slide over this level plain of ice is equally inconceivable, in view of the property of regelation of ice under pressure. Owing to this property the glacier and the lake ice would become one mass, and would move on together under the law of decreasing velocity with depth. This, however, is of little importance, if, as I conceive the supposition of the formation of an ice-sheet at the water level for fifty miles
1 A. Falsan, La Période, Glaciaire, p. 135, 137.
in advance of the glacier is an impossible one. The only other theory is, that the lake was filled up by alluvium before the ice age, and that the glacier re-excavated it. I have, however, already given reasons why the glacier would not have done so, and the very existence of this ancient alluvium in the course of the ancient glacier is a proof that it did not do so. This theory seems now to have no supporters.
Summary of the Evidence.—As the subject here discussed is very complex, and the argument essentially a cumulative one, it will be well briefly to summarize its main points.
In the first place, it has been shown that the valley-lakes of highly glaciated districts form a distinct class, which are highly characteristic if not altogether peculiar since in none of the mountain ranges of the tropics or of non-glaciated regions over the whole world are any similar lakes to be found.
The special conditions favourable to the erosion of lake-basins and the mode of action of the ice-tool are then discussed, and it is shown that these conditions have been either overlooked or ignored by the opponents of the theory of ice-erosion.
The objections of modern writers are then considered, and they are shown to be founded either on mistaken ideas as to the mode of erosion by glaciers, or on not taking into account results of glacier-action which they themselves either admit or have not attempted to disprove.
The alternative theory — that earth-movements of various kinds led to the production of lake-basins in all mountain ranges, and that those in glaciated regions were preserved by being filled with ice—is shown to be beset with numerous difficulties, physical, geological, and geographical, which its supporters have not attempted to overcome. It is also pointed out that this theory in no way explains the occurrence of the largest and deepest lakes in the largest river valleys, or in those valleys where there was the greatest concentration of glaciers, a peculiarity of their distribution which points directly and unmistakably to ice-erosion.
A crucial test of the two theories is then suggested and
it is shown that both the sub-aqueous contours of the lake-basins, and the superficial outlines of the lakes, are exactly such as would be produced by ice-erosion, while they could not possibly have been caused by submergence due to any form of earth-movements. It is submitted that we have here a positive criterion, now adduced for the first time, which is absolutely fatal to any theory of submersion.
Lastly, the special case of the Lake of Geneva is discussed, and it is shown that the explanation put forth by the anti-glacialists is wholly unsupported by facts, and is opposed to the known laws of glacier motion. The geologists who support it themselves furnish evidence against their own theory in the ancient alluvium at Geneva on which the glacial deposits rest, and which is admitted to be mainly derived from the distant Alps. But as all alluvial matter is necessarily intercepted by large and deep lakes, the presence of this Alpine alluvium immediately beneath the glacial débris at the foot of the lake, indicates that the lake did not exist in pre-glacial times, but that the river Rhone flowed from the Alps to Geneva, carrying with it the old alluvium consisting of mud, sand, and gravel, which it had brought down from the mountains. Still more conclusive, however, is the fact that the three special features which have been shown to indicate erosion rather than submergence are present in this lake as fully as in all other Alpine valley-lakes and unmistakably point to the glacial origin of all of them.
On the whole, I venture to claim that the facts and considerations set forth in this chapter show such a number of distinct lines of evidence, all converging to establish the theory of the ice-erosion of the valley-lakes of highly glaciated regions—a theory first advocated by the late Sir Andrew Ramsay—that that theory must be held to be established, at all events provisionally, as the only one by which the whole body of the facts can be explained and harmonized.
EXPLANATION OF MAP OF THE ANCIENT RHONE GLACIER.
Outline map of part of France and Switzerland, showing the course of the ancient Rhone glacier at the period of its greatest extension, to illustrate the account here given of the dispersal of Alpine erratics, as well as of the erosion of the basin of the Lake of Geneva by the glacier.
EXPLANATION.
1. The parallel and diverging blue lines show the courses of the various streams of erratics carried by the glacier.
2. The arrows at end of these lines show the positions of the most distant Alpine erratics, or of drift containing fragments of Alpine rocks.
3. The Blue figures show the height in feet of the ice-surface above sea-level, as shown by ice-worn rocks or erratics in the vicinity. The actual elevation of the ice was perhaps much greater.
4. The Black figures show the heights of land or water surfaces above sea-level.
5. The tributary glaciers (except that of the Arve) are omitted for clearness; but the whole area between the Alps and the Jura, except a few high summits, was buried in ice.
N.B. The lines showing the course of the erratics are taken from A. Falsan's large map published in 1887.
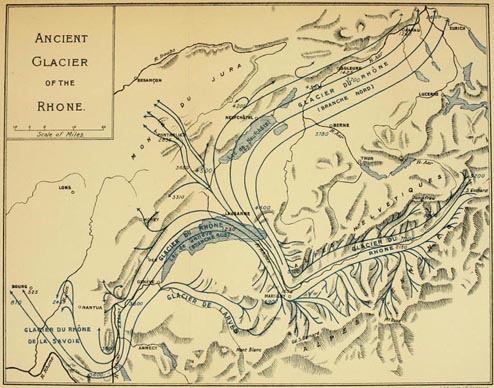
CHAPTER VI
THE GORGE OF THE AAR AND ITS TEACHINGS
THERE is perhaps no valley in Switzerland that offers to the tourist so much variety and grandeur, and to the glacialist so much instruction, as the Haslithal or valley of the Aar. I visited it for the first time last summer, walking over the Grimsel Pass to the Hospice and the Aar glacier, and thence along the old mule-track and fine new carriage-road to Meiringen; staying there three days to visit the Reichenbach Falls, the Kirchet Hill, the gorge of the Aar, and other interesting localities. It seemed to me at the time that the phenomena presented by this valley afforded a striking example of the vast amount of glacial erosion, and that some of the conclusions to which they point had been overlooked by English writers. They give us, in fact, a fresh and very powerful argument in support of the power of the ancient glaciers both to deepen valleys and to grind out lake-basins; and I now propose to lay before my readers the facts which seem to me to prove the correctness of this view.
The Grimsel Pass is a low one, only a little over 7,000 feet, but for this reason, and because it lies directly between extensive areas of perpetual snow, which give rise to some of the finest glaciers in Switzerland, it has been very largely ice-ground and presents a scene of savage grandeur which is often absent from higher passes. Everywhere the rocks are ground into huge domes or smooth slopes or rounded hollows, and these ice-ground
VOL. I. K
contours extend to at least a thousand feet higher, above which level the mountains rise in sharp peaks or serrated ridges. The descent towards the Grimsel Hospice is very grand, owing to the enormous surfaces of smooth ice-ground rocks of the hardest gneiss, which plunge down at a very high angle for nearly a thousand feet into the curious little enclosed valley, with its two small rock-basin lakes, in which the hospice is situated. Here we see an example of the effects of a kind of eddy in the old ice streams to which I think sufficient attention has not been paid. The torrent from the Aar glacier comes in from the west, but before reaching the Hospice turns off abruptly through a narrow gorge into the main valley, running at first nearly north. But looked at from above, this gorge is invisible, and it seems as if the valley from the glacier continued through the two small lakes further to the east. It is evident that when this district was buried deep in ice very little of it could escape through the gorge, but must have flowed over the higher slopes, while the portion in the valley, fed by ice-streams from nearly opposite directions, would acquire a slow eddying motion which would greatly aid its grinding power, and thus account for the land-locked valley and the two small rock-basin lakes.
Proceeding down the valley we see on all sides precipitous slopes of ice-worn rocks, some of which are so smooth and so extensive that steps have had to be cut in them to form the old mule-path, the new road here going on the opposite side of the valley. Usually the valley is narrow or V-shaped, but in several places, where it widens out, as a little below the Grimsel gorge, above the Handeck fall, and especially at Guttannen about half way to Meiringen, there is more or less of flat valley bottom, suggesting filled-up lake-basins, and showing that in these places the valley is of the U-form, which is held to be the characteristic result of glacial erosion. Why this form is not more general seems to me to be due to the character of the pre-existing valleys. Where these were narrow and precipitous, with the features of ordinary mountain gorges, the greater part of the weight of the ice would rest upon their slopes, which would be ground or split off as the ice
moved downwards, while it would hardly penetrate at all into the narrow valley bottom, or if it did, would be so checked in its motion by the friction of the rugged sides, craggy projections, and constant change of direction as to cause very little erosion. For this reason the higher narrow and deep valleys exhibit only those indications of superficial grinding which have been thought to prove that glaciers, however large, have no great erosive power; but whenever we reach the lower levels, where the original valleys were themselves wider, we find proofs of much greater grinding power, both vertical and horizontal, leading to the characteristic U-forms, which are so well seen in the valley of Meiringen and in the lakes below it, in the Lauterbrunnen valley in the Rhone valley between Martigny and Brieg, and especially in the grand vertical precipices which bound the Lake of Lucerne throughout its eastern half and its south-western arm.
It is, however, after passing through the narrow gorge between the hamlets of Inner and Outer Urweid, with its picturesque waterfall, which the coach road passes by tunnelling under it, that we approach the most remarkable feature of the district. The valley rapidly widens, with a perfectly flat bottom, till at Innertkirchen it becomes nearly a mile wide. Here, on the right, the united waters of the Gadmen and Genthal valleys form a junction with the Aar, issuing out of a narrow gorge between lofty rocks; but the most striking object is the barrier of the Kirchet, which extends quite across the valley, rising abruptly to the height of 500 feet above it, and appearing entirely to dam the course of the foaming torrent which has now become a good-sized river. Passing along the high road, which by three great zigzags mounts up the steep ascent, the entrance to the gorge is not visible; and it is only after crossing the hill and descending on the other side into the wide alluvial plain of Meiringen that the stream is again seen as a considerable and rapidly flowing river. It has passed through the hill by the celebrated Aarschlucht, one of the most remarkable gorges in Europe, which is now rendered easily accessible by a wooden platform suspended a few feet above the surface of
K 2
the water, and extending for about two-thirds of the total length of the gorge, or nearly half a mile.
On entering the gorge we are struck by its extreme narrowness, usually not more than six or eight feet, often

FIG. 23.—THE AARSCHLUCHT.
not more than four, and in some places even less; its great depth, from 200 to 300 feet, and its remarkable uniformity of width not perceptibly increasing upwards. The sides are cut away in curves or hollows such as would be caused by tumultuous eddying waters; the walls fre-
quently overhang and then recede again, so that it is only here and there that they are sufficiently vertical to enable us to catch a glimpse of the sky, and wherever we do so we see that the upper edges of the chasm are little, if any, farther apart than are the rock-walls between which we walk. The whole surface of the rock—a hard crystalline limestone—is evidently water-worn, never presenting surfaces due to fracture except perhaps where a lateral stream enters by a picturesque cascade falling over a vertical rock, and where the gorge opens out so that daylight and sunlight freely enter it. The artificial causeway finishes where a dry lateral gorge, with a steeply rising floor of earth and vegetable débris, affords an exit to the plateau and the road from Meiringen. This short lateral gorge is of especial interest, because it reproduces in almost every detail the features of the main gorge, being about the same average width, having similar walls of hollow curvilinear form, and being equally narrow to the very top. This lateral gorge is, however, quite dry, and even in the wettest seasons can hardly have more than a trickle of water because it has no catchment basin, opening out as it does on the top of the bossy limestone rocks of the narrow ridge of the Kirchet. Hence we reach the important conclusion that this gorge could not have been formed by water derived from ordinary streams, unless at a period so remote that the whole surface contours of the district were very different from what they are now. The only explanation that seems to accord with the facts is, that we have here the result of the action of sub glacial torrents acting throughout the whole period during which the area was buried in ice. Thus only are we able to explain the fact of the almost uniform narrowness of the gorge from bottom to top, since during the process of its formation the rock-walls would be preserved from ordinary denuding agencies, probably by a plug of ice, and be kept at a nearly uniform temperature. Hence we have the actual surface as it was left by the glacial waters, and its extreme narrowness together with the luxuriant vegetation which covers the plateau and fringes the edge of the chasm, aided also by the comparatively mild climate of the lower valley,
have preserved for us one of the most curious and instructive products of the great ice-age. This view of the origin of the gorge is that adopted by Professor Bonney, who, in his paper read before the Royal Geographical Society in 1893, says, "This chasm has been sawn by the sub-glacial torrent, while the ice itself has moulded every rock on the barrier into billowy undulations." I may, therefore, take it for granted that this view is held by most geologists who have attended to the subject, and it appears to me to be the only reasonable one; yet it is so important, and leads to conclusions so entirely opposed to the views expressed by Professor Bonney in the same paper, that it seems advisable to ascertain whether any collateral evidence can be obtained in support of it.
There are in Switzerland a number of other gorges which have the same general characters, of being decidedly water-worn throughout, very narrow and deep, and of approximately the same width from bottom to top; but they are all formed by lateral streams where entering into a main valley, not in course of the main valley itself. Such are the gorges of the Trient at its outlet to the Rhone, that of Pfœffers on the Rhine, and many others of less importance on the tributaries of other Alpine rivers. In many of the larger valleys almost every lateral tributary enters the main stream either by a cascade or by a gorge of this character, these gorges usually being the outlet of the drainage of considerable valleys, the stream appearing to have cut through a rocky barrier similar to that of the Aar, but on a less impressive scale. Now all the gorges which have these special characters have almost certainly been formed in the same way—by sub-glacial torrents, and, if so, the presence of such gorges would be an indication of geologically recent glaciation. It is true that similar gorges may have been sometimes formed, without the intervention of an ice-covering, in rainless districts where sub-aerial denudation would not eat away the upper lips of the chasm as it was being formed. This is to some extent the case in the cañons of the Colorado; but although the lower portions may exhibit the effects of water erosion only, being by their great depth kept at a nearly uniform
temperature, the upper portions of these cañons have been greatly widened and everywhere exhibit surfaces of fracture and sub-aerial erosion, largely due to the great and rapid alternations of temperature to which they are exposed.
If this view of the formation of these water-worn gorges is correct they ought to be found only in regions which have been recently glaciated, and it will be interesting to inquire if this is the case. In our own country we have many small gorges of this character, in Wales, the Lake District, and Scotland, that of Dungeon Gill, in Westmoreland, being an example; but none are to be found in decidedly non-glaciated areas, such as Devonshire, though narrow ravines are common enough. So in the Northern United States there are many such gorges, the Ausable Chasm in the Adirondacks, and Watkin Glen near Seneca Lake are well-known tourist resorts; but in the Southern States, beyond the glaciated area, there are no similar gorges, although the southern Alleghanies are loftier than farther north, and contain much grand and picturesque mountain scenery and many waterfalls and deep ravines, but these are all of the rugged and weathered type.
The best test, however, of these vertical gorges being really produced by sub-glacial torrents, will be to ascertain whether they occur in any extensive tropical mountain-region where there has certainly been no recent glaciation. Brazil seemed to me to be such a country, of great extent, mountainous throughout a large part of its area, and very thoroughly explored. I therefore wrote to my friend, Professor Branner, of the Stanford University, California, who spent three years in making a geological survey of Brazil, visiting every part of the country and paying special attention to all the physical features of the surface.
In reply to my inquiry, Professor Branner writes as follows:—
"What you say of certain channels in Switzerland is certainly most interesting…I recall the fact that there are just such gorges at Grindelwald and at Zermatt opening from beneath the glaciers. I have turned over in my
mind all the facts I have in regard to stream-channels in unglaciated areas, and I cannot recall a single instance of such a gorge. Those most nearly approaching it are some of the mountain gorges in the diamond regions of Brazil, but those belong to an entirely different type." (Sketches are given showing deep valleys vertical on one side and sloping on the other.) "There are no such gorges as you describe in any part of Brazil that I have seen. In the United States there are such in the glaciated area, such as Watkin Glen in the State of New York. South of the glaciated area I have no recollection of ever having seen such gorges."
By the kindness of Mr. Percy Kendall I am able to give a photo-plate of one of the gorges referred to by Professor Branner, that of the Gorner glacier at Zermatt. It exhibits the plug of rotten ice filling the narrow gorge and the sub-glacial torrent rushing out beneath it. It thus shows us on a very small scale the actual mode of formation of the Aarschlucht and other Alpine gorges of the same character.
I have endeavoured, by means of collateral evidence, to support the generally accepted view that the gorge of the Aar and others of like nature have been wholly formed by sub-glacial torrents, because, if they have been so formed, they afford a distinct class of evidence in favour of the large amount of glacial erosion in general, and of the theory of the glacial origin of the Swiss valley-lakes in particular, but which has, so far as I know, been overlooked by most writers on the subject. The nature and bearing of this evidence I will now briefly discuss.
The large-scale Swiss Government Map shows us that the valley above the gorge of the Aar is less than 20 feet above the level of the valley below the gorge. Both are flat alluvial plains having every appearance of being filled-up lake basins. The Kirchet hill, pierced by the gorge, is about 450 feet high, the highest ice-ground bosses being nearly 600 feet, while the lowest point over which the road passes is a little more than 300 feet, but this notch is apparently the continuation of the lateral gorge already described. During the long time occupied

FIG. 24.—TERMINAL GORGE OF THE GORNER GLACIER, ZERMATT.
in the cutting of the great gorge by the sub-glacial torrent, the Kirchet itself must have been greatly lowered, and was probably, when the ice first reached it, at least 150 feet higher than its present highest bosses, or about 800 feet above the present level of the upper plain. What is the depth of the alluvial deposit in this plain we do not know, but as it extends more than two miles up the valley, with a rise of about 30 feet in a mile, it is probably not less than 200 feet deep. But if we go back to the pre-glacial period before the gorge was cut, the valley-bottom must have sloped upward from the level of the Kirchet, and must therefore have been about 800 feet higher than it is now. It follows, that while the gorge was being cut, the floor of the valley above it was being ice-ground, resulting in a basin about 1,000 feet deep if we add 200 feet for the supposed depth of alluvial deposits.
The cause of the exceptional grinding power of the glacier in this part is very clear. From the Grimsel downwards all the lateral tributaries are short, but just where the valley widens above Innertkirchen, the Urbach valley opens from the south, bringing the outflow of the Gauli glacier, and a little farther on a large valley opens from the north-east, having a drainage area about equal to that of the Aar itself, and which must have nearly doubled the size of the main ice-stream by the tributaries from the Trift, the Susten, and the Titlis glaciers. The effect of these great inflows from different directions must have been to cause a heaping up of the ice, and to give it an eddying motion,1 thus producing the powerful grinding tool which hollowed out the rock-basin above the Kirchet. The influx of this great ice-stream from the north-east will also explain the curious abruptness of the Kirchet hill, which, almost like a wall, blocks up the valley, and
1 Some writers object that there is no evidence of eddies occurring in ice. But Mr. E. J. Garwood, in his paper on the Glacial Phenomena of Spitzbergen (Q.J.G.S., Nov. 4, 1899, p. 683), actually observed eddies in the glacier. He says: "In places also eddies may be observed distinctly comparable with the swirl of river water in the neighbourhood of submerged rocks, so that, even where no rock is visible, the presence of buried mountain ridges may often be inferred."
which Professor Bonney has advanced as an argument against the theory I am advocating. He says: "This grassy basin might well be claimed as an instance of glacial erosion—by which indeed it possibly may have been deepened; but if we attribute to this agency the removal of all the rock between the summit level of the barrier and the present meadow-floor, how are we to explain the existence of the steep rocky slope down which the road to Imhof descends in zigzags? The slopes, in the teeth of an advancing glacier are always comparatively gentle, and very unlike those which are presented by this rocky rib." This would be a valid objection if the Aar glacier had continued its course in a straight, or nearly straight, line to Meiringen; but the influx of a large glacier-stream from the north-east must have so diverted that of the Aar, that the resultant flow would have been across the lower valley and almost along the steep face of Kirchet instead of directly across it. This would have been the case, because the glacier stream from the north-east was not only equal in size to that of the Aar valley, but had a more rapid descent, and, therefore, a quicker flow. In the last five miles the Aar valley has a fall of about 1,500 feet, while the two northeastern valleys have an average fall of about 2,000 feet; and they are also much wider, which would still further facilitate rapidity of outflow.
The influx of this powerful glacier stream from the northeast well explains the main physical features of the district, especially the nearly uniform plateau of the Geissholz Alp, at about the same level as the top of the Kirchet, bounded by the line of precipices extending to the Reichenbach falls, and continued along the south side of the valley. On the north side the plateaux are at a higher level, indicating that the motion of the glacier was there less rapid, and its grinding power correspondingly diminished. While the main body of the glacier thus moved onward towards Thun, the meeting of three glacier streams, of unequal size and velocity and from different directions, would produce a vast ice-eddy at their junction,
accompanied by a heaping up of the surface and great grinding power, thus leading to those curiously embayed hollows around Eppigen and Imhof, which are not found on the opposite side of the valley, where the glacier had a direct onward motion.
It appears, therefore, that the singular phenomena of a great valley barred across by a precipitous rocky ridge, which is pierced only by a narrow water-worn gorge, admittedly sawn down by the débris-laden water of the sub-glacial torrent, does afford a most striking additional proof of the power of the old glaciers to grind out rock-basins. The only escape from this conclusion is to call in the aid of hypothetical local subsidences or elevations of which no direct evidence has yet been found. And when we see that, besides all the existing valley-lakes of glaciated countries, we have also to account for these gorge-pierced rocky dams with the filled-up lake-basins above them, and for the number of other filled-up rock-basins which occur in the course of all the great Alpine valleys, the explanation by subsidence becomes more and more difficult and improbable. And the difficulty is still further increased by the consideration, that all these earth-movements must have been comparatively recent, that in all cases they must have been so rapid that the erosion of the valley did not keep pace with them, and that, in all non-glaciated areas, either no earth-movements have occurred within the recent period, or, if they have occurred, they have failed, in every single case, to produce a lake-basin, because erosion has fully kept pace with elevation. A theory which involves such a series of improbable assumptions, and which is not supported by any direct evidence, stands self-condemned.
But besides this body of evidence, in itself almost conclusive, I have adduced in the preceding chapter a new and independent argument founded on the characteristic contours of all these valley-lakes, which were shown to be such as could be produced only by ice-erosion in a pre-existing valley, while any form of damming up by subsidence above or elevation below the lake would necessarily result in a very different contour.
I refer to this argument, because, since my articles first appeared, I have received from two American geologists copies of papers showing that similar features have been observed in some of the American valley-lakes, and the same general conclusion has been drawn from them. In a paper on "Glaciation in the Finger Lake region of New York," by Dr. D. F. Lincoln (in the American Journal of Science, vol. xliv., October, 1892), it is shown that Seneca Lake is bordered through a great part of its circumference by vertical cliffs of rock, that it undulates in moderate curves, and that inflowing streams with rocky beds enter it, either near the level of the water or at a considerable elevation. In some cases a lateral stream flows over nearly level ground to the cliff-edge without any valley banks, suggesting that the main valley occupied by the lake was formerly at this high level. The conclusion is reached that this and other lakes with similar characteristics have been "deeply gouged by ice." In a later paper (dated August, 1893) he notices the absence of side-valleys along Seneca and Cayuga Lakes, and points out the remarkable difference in this respect between these lakes and valleys situated south of the great moraine, which latter are "notched down to the flood-plain level every mile or two," while "the lake-side for stretches of ten to fifteen miles seems like an artifically smoothed garden slope."
In the Bulletin of the Geological Society of America, vol. v., pp. 339–356 (March, 1894), Mr. Ralph S. Tarr has a paper on "Lake Cayuga a Rock-basin," illustrated by a map and section. He maintains that Lake Cayuga is situated in a rock-basin, about 435 feet deep, and formed by glacial erosion; the proof being that the pre-glacial tributaries of the valley are rock-enclosed, and that their lowest points are above the present lake-surface. The map shows all the lakes of this region to have the very same kind of contour as that on which I founded my argument.
I venture to hope that the brief sketch I have here given of one of the grandest and most interesting valleys
in Switzerland and its bearing on that most curious and fascinating problem—the Great Ice Age, its causes, its duration, and its effects, may attract the attention of some readers who are about to visit the district, and enable them, when they come in sight of the Kirchet or visit the celebrated gorge, to appreciate the remarkable and almost unique character of the natural phenomena before them, and the marvellous episode in the recent geological history of the north-temperate zone which has produced them.
APPENDIX
An American critic of this article, in Science (N. S., vol. iv., No. 95), declares that my "derogatory references to the explanation of lakes by subsidence or deformation seems hardly candid in face of the facts reported by many Swiss geologists." And he concludes thus: "The citation of the essays by Lincoln and Tarr concerning the Finger Lakes of New York, and the silence concerning such essays as Aeppli's on Lake Zurich, give Wallace's essay an air of special pleading."
Finding it stated in Professor James Geikie's recent work on "Earth Sculpture" (1898) that he does not know of any valley-lake that has been shown to be formed by earth-movements, and as he is one of the highest authorities on everything connected with glaciers, I requested his judgment as to the special case relied on by my American critic, that of Lake Zurich, as investigated by Aeppli. His reply, which I am kindly permitted to quote, is as follows:—
"It is quite true that geologists have often stated their belief that the great Alpine lakes are due to deformation, but it is seldom that any evidence of deformation is adduced. Yet if the belief were well founded it should not be difficult, by means of maps and accurately drawn sections, to show that it is so. According to Professor Heim, the older quaternary deposits along the base of the Alps, in the region occupied by Lakes Constance and Zurich, have been affected by a flat geo-anticline (about 300—400 metres in height). If this broad fold had any effect in deforming the valley-bottoms, we should expect tectonic hollows to appear in these just above (i.e., higher up the valleys than) the flat swelling. But, as Professor Penck has pointed out, both Lake Constance and Lake Zurich extend right across the geo-anticline—the latter lake attaining its greatest depth near the back of the broad arch. That the valley of the Linth has been disturbed by crustal movement in early Pleistocene times is
shown by the fact that certain ancient terraces of erosion and deposits of river-gravel, seen on both sides of the lake, have been tilted up. From a point midway between the two extremities of the lake they gradually rise higher and higher in the direction of Zurich, until they reach their culminating point at Männedorf and Käpfnach, some nine or ten miles above Zurich. Obviously, the terraces in question are the work of the Linth, and must have been formed before the lake came into existence. But surely the simple explanation is that here, as in so many other regions, fluviatile erosion has kept pace with, or even exceeded, crustal deformation. The old river Linth has behaved just as other rivers under similar circumstances—it has cut its way across a gradually-developing deformation. Aeppli, following Heim, believes that the upper part of the lacustrine hollow (i.e., between Männedorf and the head of the lake) has been caused by a depression of the great Alpine massif, producing a reversal of the valley-slope. But since such a movement will not account for the lower section of the hollow (i.e., between Männedorf and Zurich), another movement is invoked to explain its occurrence. We are asked to believe that the portion referred to is due to flexing and folding—a continuation of the flexures of the Jura mountains. It seems to me just as easy to believe that the entire lacustrine hollow occupies an area of special depression! That flexing and folding have taken place cannot be doubted, but there is nothing whatever to show that the river Linth was incapable of cutting across the inequalities as fast as these were developed. Even if we could accept Heim's notion of a depression of the Alpine massif, and believe with him that the slope of the old valley was in this way reversed, we should not necessarily be able to account for the existence of Lake Zurich. Any great crustal movement of the kind, we have no reason to doubt, would be gradual—so gradual and protracted that the river would continuously fill up the slowly-deepening depression, and thus a lake could not come into existence. [In illustration, I may refer to the deep basin in the valley of the Rhine in Hesse, the bottom of which is 270 feet deeper than the lowest
part of its rim at Bingen. This basin is filled (not with lacustrine, but) with fluviatile deposits alone, showing that the process of infilling has kept pace with the movement of depression.] Lake Zurich, I would only add, is the bed of the old Linth glacier, and its dimensions are in proportion to those of that extensive ice-stream. The lacustrine hollow has obviously been excavated in the valley of the Linth long after that river had cut its way across the crustal deformations I have referred to."
The account here given of Aeppli's work shows that he adduces no evidence whatever of the existence of the alleged deformation, but only an assumption of an earth-movement sufficient to account for the formation of Lake Zurich. There is, however, the clearest evidence of another earth-movement on the very site of the lake in question, but in the opposite direction—that is, an elevation instead of a depression, through which the old river Linth cut its way as fast as the ground rose; and, subsequently, what had been the crown of the elevated ground was covered by the Linth glacier, and now forms the centre of the lake! It will, I think, be admitted that my critic was hardly justified in his accusation of "special pleading" on my part.
VOL. I. L
CHAPTER VII
MONKEYS: THEIR AFFINITIES AND DISTRIBUTION
IF the skeletons of an orang-utan and a chimpanzee be compared with that of a man, there will be found to be the most wonderful resemblance, together with a very marked diversity. Bone for bone, throughout the whole structure, will be found to agree in general form, position, and function, the only absolute differences being that the orang has nine wrist bones, whereas man and the chimpanzee have but eight; and the chimpanzee has thirteen pairs of ribs, whereas the orang, like man, has but twelve. With these two exceptions, the differences are those of shape, proportion, and direction only, though the resulting differences in the external form and motions are very considerable. The greatest of these are, that the feet of the anthropoid or man-like apes, as well as those of all monkeys, are formed like hands, with large opposable thumbs fitted to grasp the branches of trees but unsuitable for erect walking, while the true hands have weak small thumbs but very long and powerful fingers, forming a hook rather than a hand adapted for climbing up trees and suspending the whole weight from horizontal branches. The almost complete identity of the skeleton, however, and the close similarity of the muscles and of all the internal organs, have produced that striking and ludicrous resemblance to man which every one recognises in these higher apes and, in a less degree, in the whole monkey tribe; the face and features, the motions, attitudes, and gestures being often a strange caricature of humanity.
Let us, then, examine a little more closely in what the resemblance consists, and how far, and to what extent, these animals really differ from us.
Besides the face, which is wonderfully human—although the absence of any protuberant nose gives it often a curiously infantile aspect, monkeys, and especially apes, resemble us most closely in the hand and arm. The hand has well-formed fingers with nails, and the skin of the palm is lined and furrowed like our own. The thumb is, however, smaller and weaker than ours, and is not so much used in taking hold of anything. The monkey's hand is, therefore, not so well adapted as that of man for a variety of purposes, and cannot be applied with such precision in holding small objects, while it is unsuitable for performing delicate operations such as tying a knot or writing with a pen. A monkey does not take hold of a nut with its fore-finger and thumb as we do, but grasps it between the fingers and the palm in a clumsy way, just as a baby does before it has acquired the proper use of its hand. Two groups of monkeys—one in Africa and one in South America—have no thumbs on their hands, and yet they do not seem to be in any respect inferior to other kinds which possess it. In most of the American monkeys the thumb bends in the same direction as the fingers, and in none is it so perfectly opposed to the fingers as our thumbs are; and all these circumstances show that the hand of the monkey is, both structurally and functionally, a very different and very inferior organ to that of man, since it is not applied to similar purposes, nor is it capable of being so applied.
When we look at the feet of monkeys we find a still greater difference, for these have much larger and more opposable thumbs than their hands have, and are therefore more like our hands; and this is the case with all monkeys, so that even those which have no thumbs on their hands, or have them small and weak and parallel to the fingers, have always large and well-formed thumbs on their feet. It was on account of this peculiarity that the great French naturalist Cuvier named the whole group of monkeys Quadrumana, or four-handed animals, because,
L 2
besides the two hands on their fore-limbs, they have also two hands in place of feet on their hind-limbs. Modern naturalists have given up the use of this term, because they say that the hind extremities of all monkeys are really feet, only these feet are shaped like hands; but this is a point of anatomy, or rather of nomenclature, which we need not here discuss.
Let us, however, before going further, inquire into the purpose and use of this peculiarity, and we shall then see that it is simply an adaptation to the mode of life of the animals which possess it. Monkeys, as a rule, live in trees, and are especially abundant in the great tropical forests. They feed chiefly upon fruits, but occasionally eat insects and birds'-eggs, as well as young birds, all of which they find in the trees; and, as they rarely have occasion to come down to the ground, they travel from tree to tree by jumping or swinging, and thus pass the greater part of their lives entirely among the leafy branches of lofty trees. For such a mode of existence, they require to be able to move with perfect ease upon large or small branches, and to climb up rapidly from one bough to another. As they use their hands for gathering fruit and catching insects or birds, they require some means of holding on with their feet, otherwise they would be liable to continual falls, and they are able to do this by means of their long finger-like toes and large opposable thumbs, which grasp a branch almost as securely as a bird grasps its perch. The true hands, on the contrary, are used chiefly to climb with, and to swing the whole weight of the body from one branch or one tree to another, and for this purpose the fingers are very long and strong, and in many species they are further strengthened by being partially joined together, as if the skin of our fingers grew together as far as the knuckles. This shows that the separate action of the fingers, which is so important to us, is little required by monkeys, whose hand is really an organ for climbing and seizing food, while their feet are required to support them firmly in any position on the branches of trees, and for this purpose it has become modified into a large and powerful grasping hand.
Another striking difference between monkeys and men is that the former never walk with ease in an erect posture, but almost always use their arms in climbing or in walking on all-fours like most quadrupeds. The monkeys that we see in the streets dressed up and walking erect, only do so after much drilling and teaching, just as dogs may be taught to walk in the same way; and the posture is almost as unnatural to the one animal as it is to the other. The largest and most man-like of the apes—the gorilla, chimpanzee, and orang-utan—also walk usually on all-fours; but in these the arms are so long and the legs so short that the body appears half erect when walking; and they have the habit of resting on the knuckles of the hands, not on the palms like the smaller monkeys whose arms and legs are more nearly of an equal length, which tends still further to give them a semi-erect position, Yet they are never known to walk of their own accord on their hind legs only, though they can do so for short distances, and the story of their using a stick and walking erect by its help in the wild state is not true. Monkeys, then, are both four-handed and four-footed beasts; they possess four hands formed very much like our hands, and capable of picking up or holding any small object in the same manner; but they are also four-footed, because they use all four limbs for the purpose of walking, running, or climbing; and, being adapted to this double purpose, the hands want the delicacy of touch and the freedom as well as the precision of movement which ours possess. Man alone is so constructed that he walks erect with perfect ease, and has his hands free for any use to which he wishes to apply them; and this is the great and essential bodily distinction between monkeys and men.
We will now give some account of the different kinds of monkeys and the countries they inhabit.
The Different Kinds of Monkeys and the Countries they inhabit.
Monkeys are usually divided into three kinds—apes, monkeys, and baboons;but these do not include the
American monkeys, which are really more different from all those of the Old World than any of the latter are from each other. Naturalists, therefore, usually divide the whole monkey-tribe into two great families, inhabiting the Old and the New Worlds respectively; and, if we learn to remember the kind of differences by which these several groups are distinguished, we shall be able to understand something of the classification of animals, and the distinction between important and unimportant characters.
Taking first the Old World groups, they may be thus roughly defined:—apes have no tails; monkeys have tails, which are usually long; while baboons have short tails, and their faces, instead of being round and with a manlike expression as in apes and monkeys, are long and more dog-like. These differences are, however, by no means constant, and it is often difficult to tell whether an animal should be classed as an ape, a monkey, or a baboon. The Gibraltar ape, for example, though it has no tail, is really a monkey, because it has callosities, or hard pads of bare skin on which it sits, and cheek pouches in which it can stow away food; the latter character being always absent in the true apes, while both are present in most monkeys and baboons. All these animals, however, from the largest ape to the smallest monkey, have the same number of teeth as we have, and they are arranged in a similar manner, although the tusks, or canine teeth, of the males are often large, like those of a dog.
The American monkeys, on the other hand, with the exception of the Marmosets, have four additional grinding teeth (one in each jaw on either side), and none of them have either callosities or cheek pouches. They never have prominent snouts like the baboons; their nostrils are placed wide apart and open sideways on the face; the tail, though sometimes short, is never quite absent; and the thumb bends the same way as the fingers, is generally very short and weak, and is often quite wanting. We thus see that these American monkeys differ in a great number of characters from those of the Eastern hemisphere; and they have this further peculiarity, that
many of them have prehensile or grasping tails, which are never found in the monkeys of any other country. This curious organ serves the purpose of a fifth hand, and it has so much muscular power that the animal can hang by it easily with the tip curled round a branch; while it can also be used to pick up small objects with almost as much ease and exactness as an elephant's trunk. In those species which have it most perfectly formed it is very long and powerful, and the end has the under side covered with bare skin, much resembling that of the finger or palm of the hand and apparently equally sensitive. One of the common kinds of monkeys that accompany street organ-players has a prehensile tail, but not of the most perfect kind; since in this species the tail is entirely clad with hair to the tip, and seems to be used chiefly to steady the animal when sitting on a branch by being twisted round another branch near it. The statement is often erroneously made that all American monkeys have prehensile tails; but the fact is that rather less than half the known kinds have them so, the remainder having this organ either short and bushy or long and slender, but entirely without any power of grasping. All prehensile-tailed monkeys are American, but all American monkeys are not prehensile-tailed.
By remembering these characters it is easy, with a little observation, to tell whether any strange monkey comes from America or from the Old World. If it has bare seat-pads, or if when eating it fills its mouth till its cheeks swell out like little bags, we may be sure it comes from some part of Africa or Asia; while if it can curl up the end of its tail so as to take hold of anything, it is certainly American. As all the tailed monkeys of the Old World have seat-pads (or ischial callosities as they are called in scientific language), and as all the American monkeys have tails, but no seat-pads, this is the most constant external character by which to distinguish them; and having done so we can look for the other peculiarities of the American monkeys, especially the distance apart of the nostrils and their lateral position.
The whole monkey-tribe is especially tropical, only a few kinds being found in the warmer parts of the temperate zone. One inhabits the Rock of Gibraltar, and there is one very like it in Japan, and these are the two monkeys which live farthest from the equator. In the tropics they become very abundant and increase in numbers and variety as we approach the equator, where the climate is hot, moist, and equable, and where flowers, fruits, and insects are to be found throughout the year. Africa has about 55 different kinds, Asia and its islands about 60, while America has 114, or almost exactly the same as Asia and Africa together. Australia and its islands have no monkeys, nor has the great and luxuriant island of New Guinea, whose magnificent forests seem so well adapted for them. We will now give a short account of the different kinds of monkeys inhabiting each of the tropical continents.
African Monkeys.
Africa possesses two of the Simiidæ or great man-like apes—the gorilla and the chimpanzee, the former being the largest ape known, and the one which, on the whole, perhaps most resembles man, though its countenance is less human than that of the chimpanzee. Both are found in West Africa, near the equator, but they also inhabit the interior wherever there are great forests; and Dr. Schweinfurth states that the chimpanzee inhabits the country about the sources of the Shari river in 28° E. long. and 4° N. lat. The gorilla is a more recent discovery than the other man-like apes, and owing to its great size and strength and the almost impenetrable forests in which it lives, is even now but imperfectly known. It was discovered by Dr. Savage in 1847 in the region of the Gaboon river, close to the equator in West Africa. He first obtained a skull which he believed to belong to a new species of ape, and afterwards the entire animal, which he sent to America, where it was described by Professor Wyman, an anatomist of Boston. The first young gorilla was brought to Europe in 1876, but none of these apes live long in
confinement, and it is probable that no European has yet seen the full-grown living animal. It is the only large ape that spends much of its time on the ground, though it climbs well, but owing to its great weight is not so active among the higher branches as the less bulky species. The skin is black, and the hair is also black or brownish, but

FIG. 25.—THE GORILLA (Gorilla Savagei).
grey in old individuals. The wood-cut (Fig. 25) gives a good idea of its somewhat ferocious aspect, as well as its comparatively short arms, a character that distinguishes it from the chimpanzee and orang. The fingers and toes are also thicker and shorter than in the other species, in accordance with its less arboreal habits.
The other great African apes, consisting of the chimpanzee and one or two closely allied species, are much better known, though even of these full-grown animals have never been seen alive in Europe. Young ones are, however, tolerably common, and are to be found in most Zoological Gardens. The accompanying illustration of the

FIG. 26,—CHIMPANZEE (Anthropopithecus troglodytes).
common chimpanzee (Fig. 26) well exhibits the external differences between this species and the gorilla—the longer arms, the longer hand and fingers, and the very much larger ears. The chimpanzees show great intelligence and mildness of disposition, and can be taught to perform many almost human actions, such as dressing
themselves, riding a bicycle, and many others, while they even learn to count up to three or four.
The long-tailed monkeys of Africa are very numerous and varied. One group (Colobus) has no cheek-pouches and no thumb on the hand, and many of these have long soft fur of varied colours. The most numerous group are

FIG. 27.—PLUTO MONKEY (Cercopithecus Pluto).
the Guenons, belonging to the genus Cercopithecus, rather small long-tailed monkeys, very active and lively, and often having their faces curiously marked with white or black, or ornamented with whiskers or other tufts of hair, as well shown in the Pluto monkey here figured (Fig. 27). They all have large cheek-pouches and good-sized thumbs. Many of them are called green monkeys, from the greenish-
yellow tint of their fur, and most of them are well-formed pleasing animals. They are found only in tropical Africa.
The baboons (Cynocephalus) are larger, but less numerous. They resemble dogs in the general form and the length of the face or snout, but they have hands with well-developed thumbs on both the fore and hind limbs; and this, with something in the expression of the face and their habit of sitting up and using their hands in a very human fashion, at once shows that they belong to the
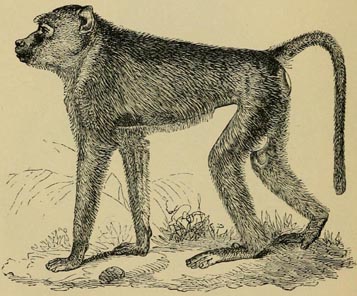
FIG. 28.—YELLOW BABOON (Cynocephalus babuino).
monkey-tribe. Many of them are very ugly, and in their wild state they are the fiercest and most dangerous of monkeys. Some have the tail very long, others of medium length, while it is sometimes reduced to a mere stump, and all have large cheek-pouches and bare seat-pads. The yellow baboon here figured is a native of West Africa, and illustrates most of the characteristic features of baboons as distinguished from other monkeys. They are found all over Africa from Egypt to the Cape of Good Hope; while one species, called the Hamadryas, extends from Abyssinia across the Red Sea into Arabia, and is the only baboon
found out of Africa. This species was known to the ancients, and it is often represented in Egyptian sculptures, while mummies of it have been found in the catacombs. The largest and most remarkable of all the baboons is the Mandrill of West Africa, whose swollen and hog-like face is ornamented with stripes of vivid blue and scarlet. This animal has a tail scarcely two inches long, while in size and strength it is not much inferior to the gorilla. These large baboons go in bands, and are said to be a match for any other animals in the African forests, and even to attack and drive away the elephants from the districts they inhabit. The baboons differ from almost all the rest of the monkey-tribe in being terrestrial rather than arboreal in their habits, though many of them can, and do, climb trees with great ease. They inhabit mostly open rocky country or among scattered trees rather than the dense forests, and their food consists of fruits and roots, as well as of reptiles, insects, and probably young ground-birds and mammals. They thus seem to be a special adaptation to the plateaus and open country of which so much of Africa consists, while the man-like apes and more active monkeys are equally well adapted to the tropical forests.
Asiatic Monkeys.
Turning now to Asia, we have first one of the best known of the large man-like apes—the Orang-utan, found only in the two large islands, Borneo and Sumatra. The name is Malay, signifying "man of the woods," and it should be pronounced órang-óotan, the accent being on the first syllable of both words. It is a very curious circumstance that, whereas the gorilla and chimpanzee are both black, like the negroes of the same country, the orang-utan is red or reddish-brown, closely resembling the colour of the Malays and Dyaks who live in the Bornean forests. Though very large and powerful, it is a harmless creature, feeding on fruit, and never attacking any other animal except in self-defence. A full-grown male orangutan is rather more than four feet high, but with a body
as large as that of a stout man, and with enormously long and powerful arms.
More is known of the habits of the Orang-utan than of any of the other large apes, which are probably not very different; a few particulars may therefore be of interest. The Orang-utan, or "Mias," as it is called in Borneo, is essentially an arboreal animal, only coming to the ground occasionally to drink or to seek juicy shoots on the banks of streams when fruit is scarce, or when desirous of crossing any opening in the forest. It lives among the tops of the lofty forest trees, on the fruits of which it feeds. The old males are often solitary, but groups consisting of a female with two or three young, and sometimes even a whole family, are seen together. Once I saw two half-grown orangs on the ground playing together. They were standing up with their arms outstretched, and holding each other as if wrestling. The moment I saw them they saw or heard me, and got up the nearest tree out of danger. This was the only time I saw them standing on their legs nearly erect. The old ones build a kind of nest, or rather a platform of branches to sleep on. Numbers of these nests are to be seen in the forests where orangs are numerous, and they probably build one in any place where fruit is abundant, and where they are likely to remain some time. The orangs move very deliberately about the trees, especially when they have to pass from one tree to another. To do this they choose a place where the branches of two trees touch or rather intermingle, and moving very carefully along the branch they have chosen and holding tight with their feet, they stretch out their long arms one after the other, binding together the twigs of the two opposing branches in their hands, and then swing themselves across till they can grasp the further branch with their feet, then they gradually move on till they get a secure hold of the other tree. This cautious procedure is often necessary in order that the slender branches may not be broken by their great weight, causing a dangerous fall.
While shooting these animals in Borneo for their skins and skeletons, I often saw them pass from tree to tree in
this way, and they generally were able to travel through the forest overhead as quickly as I could run along underneath, looking up frequently to keep them in sight. Once I saw one build a nest. I had wounded him severely and expected he would drop to the ground, but he got up as high as he could into the tree, and choosing a forked branch he stretched out his arms, broke off or cracked the smaller branches and laid them across the fork, and in a short time had made a platform which completely hid him from below. The next day he was still there but dead, as could be seen by the cloud of flies above him. He was a very fine large male, and I obtained the bones some time afterwards by paying a dollar to two Malays who climbed the tree, and tying the dried skin with a long cord let it down to the ground. It has often been stated that the orang when pursued or attacked breaks off dead branches or fruit and throws them at its pursuers, while other observers have denied that it ever does so. I have, however, seen it do so myself more than once, the creatures being evidently enraged, making a curious grunting noise and throwing down dead sticks, and on one occasion the spiny fruit of the durian, in great quantities.1
The illustration (Fig. 29, next page) represents with considerable accuracy a full-grown male Orang-utan, showing the curious dilated face formed by a ridge of the skin on each side, giving a broad flat surface varying from ten to thirteen and a half inches wide in different individuals. This peculiar character is found in no other ape. The differences between the orang and the gorilla are well shown by comparing the two figures, the former having much longer arms, longer hands and feet, and weaker thumbs, all characters which are in accordance with its more arboreal habits. An interesting peculiarity common to the three great apes—the orang utan, gorilla, and chimpanzee, but not found in any other ape or monkey, is the arrangement of the hair of the arms, which grows in opposite directions from the shoulder and
1 For fuller details see my Malay Archipelago, Chap. IV., and the late Professor Huxley's Man's Place in Nature, pp. 31—42.
wrist towards the elbow, not downwards from the shoulder to the wrist as in all other mammalia. This arrangement is no doubt connected with the habits of these creatures which clasp their hands over their heads when sleeping, and the arrangement of the hair causes the rain to run off at the elbow without wetting the skin.
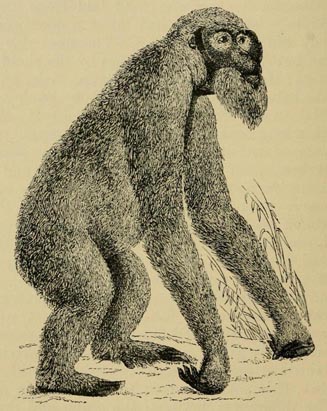
FIG. 29.—ORANG-UTAN, OLD MALE (Simia satyrus).
These great apes, like many of the smaller monkeys, are very liable to lung-disease in this country, and soon die when in confinement. This is no doubt due to the want of pure air and exercise, and it is unfortunate that none of the Zoological Societies of Europe have made suitable arrangements for giving them these essentials of health. All that would be necessary would be a rather large and lofty conservatory, containing two or three
moderate-sized trees, and opening out of this a larger enclosure containing a few clumps of loftier trees. This enclosure might be surrounded by a deep and wide trench sloping inwards, from the outer margin of the bottom of which would rise a wall or fence of a sufficient height, and perfectly smooth inside, so as to be unclimbable by these apes. Even if it were twelve feet high such a fence would offer no obstruction to the view, as the earth excavated would form a bank on the outside, reaching nearly as high as the top of the wall. Fruit trees, such as apples, crabs, damsons, cherries, chestnuts, &c, would be suitable as furnishing both food and exercise; and on mild sunny days throughout the year the orangs or chimpanzees would spend most of their time out of doors, only coming into the conservatory in very cold or wet weather. There seems no reason why, under these more favourable conditions they should not live to a fair old age. Each species should be kept separate, and the experiment should be begun with two or three young animals, whose association would be another factor tending to health. The opportunities for observation of the whole life-habits of these singular animals would be of the greatest interest, and the experiment would probably pay its cost as a public exhibition. In Ceylon, Australia, or other warm countries only an enclosed patch of woodland would be needed surrounded by such an unclimbable fence, and with a few shelters in which the animals might be fed; but even in England I feel sure that, with an arrangement such as I have suggested, these most remarkable and interesting creatures might be kept alive for many years, and might even breed in confinement.
The Gibbons.
Another group of true apes inhabit Asia and the larger Asiatic islands, and are in some respects the most remarkable of the whole family. These are the Gibbons, or long-armed apes forming the genus Hylobates, which are generally of small size and of a gentle disposition, but possessing the most wonderful agility. In these creatures the arms are as long as the body and legs together, and are so powerful that a gibbon will hang for hours sus-
VOL. I. M
pended from a branch as if this was a position of rest, or swing to-and-fro and then throw itself a great distance through the air. The arms, in fact, completely take the place of the legs for travelling. Instead of jumping from bough to bough and running on the branches, like other apes and monkeys, the gibbons move along while hanging suspended in the air, stretching their arms from bough
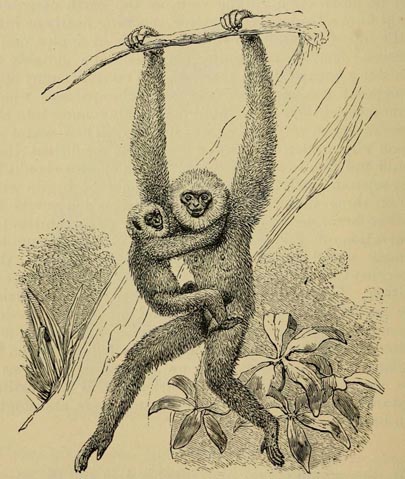
FIG. 30.—GIBBON (Hylobates entelloides).
to bough, and thus going hand over hand, as a very active sailor will climb along a rope. The strength of their arms is, however, so prodigious, and their hold so sure, that they often loose one hand before they have caught a bough with the other, thus seeming almost to fly through the air by a series of swinging leaps: and they
travel among the network of interlacing boughs a hundred feet above the earth with as much ease and certainty as we walk or run upon level ground, and with even greater speed. The woodcut of one of these long-armed apes (Fig. 30) shows a female carrying a half-grown young one, yet swinging in its favourite position.
These little animals scarcely ever come down to the ground of their own accord; but when obliged to do so they run along almost erect, with their long arms swinging round and round, as if trying to find some branch or other object to climb upon. They are the only apes which naturally walk on the ground without resting upon their hands as well as their feet; but this does not make them more like men, for it is evident that the attitude is not an easy one, and is only adopted because the arms are habitually used to swing by, and are therefore normally held upwards instead of downwards, as they must be when walking on them.
True Monkeys—Asiatic.
The tailed monkeys of Asia consist of two groups, the first of which—Semnopithecus—have no cheek pouches, but always have very long tails. They are true forest-monkeys, very active, and of a shy disposition. The most remarkable of these is the long-nosed monkey of Borneo, which is very large, of a pale brown colour, and distinguished by possessing a long, pointed, fleshy nose, totally unlike that of all other monkeys. Another interesting species is the black and white entellus monkey of India, called "Hanuman" by the Hindoos, and considered sacred by them. These animals are petted and fed, and at some of the temples numbers of them come every day for the food which the priests, as well as the people, provide for them. Monkeys of this genus are abundant from the Himalaya and north-western China to Borneo and Java, varying considerably in size and colour, but being very much alike in all essential points of structure. They may be said to correspond to the guenons of Africa, and like them are very active animals. I was often able to observe their gambols in the forests; and on one occasion, when staying for a few days at one of the travellers' rest-
M 2
houses in Sumatra, surrounded by dense forests and with no European residents within many miles, a large family of the monkeys here figured (Fig. 31) were in a tree close to the house. They took no notice of the natives who were about, but the moment I came out they showed signs of alarm, stared at me for perhaps half a minute, and then began to move off. The next tree happened to be a much lower one, and the old monkeys at once leaped on to the mass of foliage perhaps twenty or thirty feet below them. The younger ones followed, but with evident hesitation, and the last two did not like it at all, and ran a little to
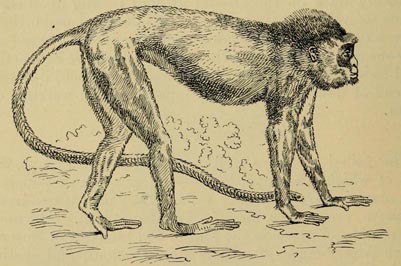
FIG. 31.—Semnopithecus melalophos.
one side or another, looking as if equally afraid to stay or go on. At length, just as the others were disappearing in the deep forest, and seeing no other means of escape, they took the dreadful jump, whirling about in the air and disappearing with a crash in the mass of foliage, but just saving themselves from coming to the ground.
The last group of Eastern monkeys are the Macaques (genus Macacus), which are more like baboons in general form, and often run upon the ground. They are more bold and vicious than the others. All have cheek pouches, and though some have long tails in others the tail is short, or reduced to a mere stump. In some few this stump is so very short that there is no external tail,

FIG. 32.—Macacus leoninus.
as in the magot of North Africa and Gibraltar, and in an allied species that inhabits Japan. The species here figured has a short tail and is a native of Arracan.
American Monkeys.
The monkeys which inhabit America form three very distinct groups: 1st—the Sapajous, which have prehensile or grasping tails; 2nd—the Sagouins, which have
ordinary tails, either long or short; and, 3rd—the Marmosets, very small creatures, with sharp claws, long tails, which are not prehensile, and a smaller number of teeth than all other American monkeys. Each of these three groups contain several sub-groups, or genera, which often differ remarkably from each other, and from all the monkeys of the Old World.
We will begin with the Howling Monkeys (genus Mycetes), which are the largest found in America, and are celebrated for the loud voice of the males. Often in the great forests of the Amazon or Oronooko a tremendous noise is heard in the night or early morning, as if
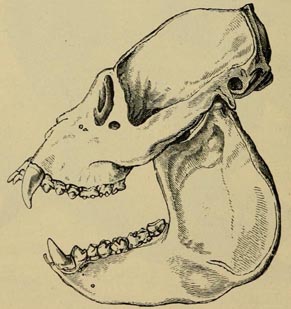
FIG. 33.—SKULL OF HOWLER (Mycetes seniculus).
a great assemblage of wild beasts were all roaring and screaming together. The noise may be heard for miles, and it is louder and more piercing than that of any other animals, yet it is all produced by a single male howler sitting on the branches of some lofty tree. They are enabled to make this extraordinary noise by means of an organ that is possessed by no other animal. The lower jaw is unusually deep, as shown in the accompanying figure, and this makes room for a hollow bony vessel about the size of a large walnut, situated under the root
of the tongue and being an outgrowth of the hyoid bone, with an opening into the windpipe by which the animal can force air into it. This increases the power of its voice, acting something like the hollow case of a violin, and producing those marvellous rolling and reverberating sounds which caused the celebrated traveller Waterton to declare that they were such as might have had their origin in the infernal regions. The howlers are large and stout-bodied monkeys with bearded faces, and with very strong and powerfully grasping tails. They inhabit the wildest forests; they are very shy, and are seldom taken captive, though they are less active than many other American monkeys.
Next come the Spider-Monkeys (genus Ateles), so called from their slender bodies and enormously long limbs and tail. In these monkeys, the tail is so long, strong, and perfect, that it completely takes the place of a fifth hand. By twisting the end of it round a branch the animal can swing freely in the air with complete safety; and this gives them a wonderful power of climbing and passing from tree to tree, because the distance they can stretch is that of the tail, body, and arm added together, and these are all unusually long. They can also swing themselves through the air for great distances, and are thus able to pass rapidly from tree to tree without ever descending to the ground, just like the gibbons in the Malayan forests. Although capable of feats of wonderful agility, when needful, the spider-monkeys are usually slow and deliberate in their motions, and have a timid, melancholy expression, very different from that of most monkeys. Their hands are very long, but have only four fingers, being adapted for hanging on to branches rather than for getting hold of small objects. It is said that when they have to cross a river the trees on the opposite banks of which do not approach near enough for a leap, several of them form a chain, one hanging by its tail from a lofty overhanging branch and seizing hold of the tail of the one below it, then gradually swinging themselves backwards and forwards till the lower one is able to seize hold of a branch on the opposite side. He then climbs
up the tree, and, when sufficiently high, the first one lets go, and the swing either carries him across to a bough on the opposite side or he climbs up over his companions. The species here figured inhabits Mexico, and exhibits the chief characteristics of the genus to which it belongs.

FIG. 34.—BLACK-HANDED SPIDER-MONKEY (Ateles melanochir).
Closely allied to the last are the Woolly Monkeys of the genus Lagothrix, which have an equally well-developed prehensile tail, but better proportioned limbs, and a thick woolly fur of a uniform grey or brownish colour. They have well-formed fingers and thumbs, both on the hands
and feet, and are rather deliberate in their motions, and exceedingly tame and affectionate in captivity. They are great eaters, and are usually very fat. They are found only in the far interior of the Amazon valley, and, having a delicate constitution, seldom live long in Europe. These monkeys are not so fond of swinging themselves about by their tails as are the spider monkeys, and offer more

FIG. 35.—HUMBOLDT'S WOOLLY MONKEY (Lagothrix Humboldtii).
opportunities of observing how completely this organ takes the place of the fifth hand. When walking about a house or on the deck of a ship the partially curled tail is carried in a horizontal position on the ground, and the moment it touches anything it twists round it and brings it forward, when, if eatable, it is at once appropriated; and when fastened up the animal will obtain any food that may be out of reach of its hands with the greatest facility,
picking up small bits of biscuit, nuts, &c., much as an elephant does with the tip of his trunk. The species here figured is a native of the Upper Amazon, and as I had one alive for some weeks I had a good opportunity of observing its habits, as above described. (Fig. 35 on preceding page.)
We now come to a group of monkeys whose prehensile tail is of a less perfect character, since it is covered with

FIG. 36.—WHITE-CHEEKED SAPAJOU (Cebus lunatus).
hair to the tip, and is of no use to pick up objects. It can, however, curl round a branch, and serves to steady the animal while sitting or feeding, but is never used to hang and swing by in the manner so common with the spider monkeys and their allies. These are rather small sized animals, with round heads and with moderately long tails. They are very active and intelligent, their limbs are not so long as in the preceding group, and though they have five
fingers on each hand and foot, the hands have weak and hardly opposable thumbs. Some species of these monkeys are often carried about by itinerant organ-men, and are taught to walk erect and perform many amusing tricks. They form the genus Cebus of naturalists, and the woodcut (Fig. 36) shows the chief characteristics of the genus, the species of which are rather numerous.
The remainder of the American monkeys have non-prehensile tails, like those of the monkeys of the Eastern hemisphere; but they consist of several distinct groups, and differ very much in appearance and habits. First we have the Sakis (Pithecia), which have a bushy tail and usually very long and thick hair, something like that of a bear. This is the case in the hairy Saki or Parauaçu of the Upper Amazon (see the lower figure in the Plate, p. 172). Mr. Bates describes it as being a timid, inoffensive creature, with a long bear-like coat of speckled grey hair. The long hair of the head hangs down, so as to partly conceal the pleasing diminutive face. It is a very delicate animal, rarely living in captivity more than a month or two even in its native country. It is found on the northern shore of the Upper Amazon as far as Peru, and a different but closely allied species is found south of the great river, which, as well as the Rio Negro, limits the range of several species both of quadrupeds and birds. In some species the tail is very short, appearing like a rounded tuft of hair; many of the species have fine bushy whiskers, which meet under the chin, and appear as if they had been dressed and trimmed by a barber, and the head is often covered with thick curly hair, looking like a wig.
The Uacari monkeys, forming the genus Brachiurus, have the face quite red, and one has the head nearly bald, a most remarkable peculiarity among monkeys. This latter species was met with by Mr. Bates on the Upper Amazon, and is figured at the top of the Plate. He describes the face as being of a vivid scarlet, the body clothed from neck to tail with very long, straight, and shining white hair, while the head is nearly bald, owing to the very short crop of thin grey hairs. As a finish to their striking physiognomy these monkeys have bushy whiskers of a sandy colour meeting under the chin, and yellowish-grey eyes.

FIG. 37.—HAIRY SAKI (Pithecia hirsuta), (lower fig.).
FIG. 38.—BALD UACARI (Brachiurus calvus), (upper fig.).
The colour of the face is so vivid that it looks as if covered with a thick coat of bright scarlet paint. These creatures are very delicate, and have never reached Europe alive.
An allied group consists of the elegant Squirrel-Monkeys (Callithrix and Chrysothrix), with long, straight, hairy tails, and often adorned with prettily variegated colours. They

FIG 39.—LEMURINE NIGHT MONKEY (Nyctipithecus lemurinus).
are usually small animals; some have the face marked with black and white, others have curious whiskers, and their nails are rather sharp and claw-like. They have large round heads, and their fur is more glossy and smooth than in most other American monkeys, so that they more resemble some of the smaller monkeys of Africa. These little creatures are very active, running
about the trees like squirrels, and feeding largely on insects as well as on fruit.
Closely allied to these are the small group of Night-Monkeys (Nyctipithecus), which have large eyes, and a round face surrounded by a kind of ruff of whitish fur, so as to give it an owl-like appearance, whence they are sometimes called owl-faced monkeys. They are covered with soft grey fur, like that of a rabbit, and sleep all day long concealed in hollow trees. The face is also marked with white patches and stripes, giving it a rather carnivorous or cat-like aspect, which, perhaps, serves as a protection, by causing the defenceless creature to be taken for an arboreal tiger-cat or some such beast of prey. The cut (Fig 39 on the preceding page) well represents the peculiarities of these remarkable monkeys.
This finishes the series of such of the American monkeys as have a larger number of teeth than those of the Old World. But there is another group, the Marmosets, forming the sub-family Hapalidæ, which have the same number of teeth as Eastern monkeys, but differently distributed in the jaws, a pre-molar being substituted for a molar tooth. In other particulars they resemble the rest of the American monkeys. These are very small and delicate creatures, some having a body only seven inches long. The thumb of the hands is not opposable, and instead of nails they have sharp compressed claws. These diminutive monkeys have long non-prehensile tails, and they have a silky fur often of varied and beautiful colour. Some are striped with grey and white, or are of rich brown or golden brown tints, varied by having the head or shoulders white or black, while in many there are crests, frills, manes, or long ear tufts, adding greatly to their variety and beauty. The species figured (on the opposite page) is of beautiful yellow and white tints, and is found in the valley of the Madeira, one of the great southern tributaries of the Amazon. The little marmosets are timid and restless; their motions are more like those of a squirrel than a monkey. Their sharp claws enable them to run quickly along the branches, but they seldom leap from bough
to bough like the larger monkeys. They live on fruits and insects, but are much afraid of wasps, which they are said to recognise even in a picture.
This completes our meagre sketch of the American monkeys, and we see that, although they possess no such remarkable forms as the gorilla or the baboons, yet they exhibit a wonderful diversity of external characters, con-
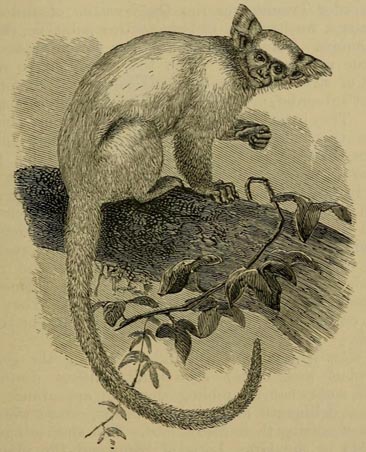
FIG. 40.—GOLDEN MARMOSET (Midas chrysoleucas).
sidering that all seem equally adapted to a purely arboreal life. In the howlers we have a specially developed voice-organ, which is altogether peculiar; in the spider-monkeys we find the adaptation to active motion among the topmost branches of the forest trees carried to an extreme point of development; while the singular nocturnal monkeys, the
active squirrel-monkeys, and the exquisite little marmosets, show how distinct are the forms under which the same general type may be exhibited, and in how many varied ways existence may be sustained under almost identical conditions.
Lemurs.
In the general term monkeys, considered as equivalent to the order Primates, or the Quadrumana of the older naturalists, we have to include another sub-type, that of the Lemurs. These animals are of a lower grade than the true monkeys, from which they differ in so many points of structure that they are considered to form a distinct sub-order, or, by some naturalists, even a separate order. They have usually a larger head and more pointed muzzle than monkeys; they vary considerably in the number, form, and arrangement of the teeth; their thumbs are always well-developed, but their fingers vary much in size and length; their tails are usually long, but several species have no tail whatever, and they are clothed with a more or less woolly fur, often prettily variegated with white and black. They inhabit the deep forests of Africa, Madagascar, and Southern Asia, and are more sluggish in their movements than true monkeys, most of them being of nocturnal or crepuscular habits. They feed largely on insects, eating also fruits and the eggs or young of birds.
The slow lemurs of South India (forming the genera Nycticebus and Loris) are small, tailless nocturnal animals, somewhat resembling sloths in appearance, and almost as deliberate in their movements, except when in the act of seizing their insect food. The species figured (Fig. 41) is the Gray Loris of Siam and the adjacent countries. It lives in the deep forests of the mountains, sleeping during the day in holes of trees, but at night roaming about among the branches in search of fruit and tender leaves, as well as insects, mice, or small birds. It moves slowly and noiselessly till it can suddenly seize its prey with its grasping hand. The grey fur renders it almost invisible at night, while its largely developed eyes
enable it to discover its prey. The Tarsier (genus Tarsius) or spectre-lemur, of the Malay islands, is a small long-tailed nocturnal lemur, remarkable for the curious development of the hind feet, which have two of the toes very short and with sharp claws, while the others have nails, the third toe being exceedingly long and slender, though the thumb is very large, giving the feet a very irregular and outré appearance. Even more curious is the Aye-aye of Madagascar (Chiromys), the most remarkable of all the lemurs. This animal has very large ears and a squirrellike tail, with long spreading hair. It has large curved

FIG. 41.—GRAY LORIS (Nycticebus cinereus).
incisor teeth, which add to its squirrel-like appearance and caused the early naturalists to class it among the rodents. But its most remarkable character is found in its fore-feet or hands, the fingers of which are all very long and armed with sharp curved claws, but one of them, the second, is wonderfully slender, being not half the thickness of the others. This curious combination of characters shows that the aye-aye is a very specialized form—that is, one whose organisation has been slowly modified to fit it for a peculiar mode of life. From information received from its
VOL. I. N
native country, and from a profound study of its organization, Professor Owen believes that it is adapted for the one purpose of feeding on the larvæ of wood-boring insects. Its large feet and sharp claws enable it to cling firmly to the branches of trees in almost any position; by means of its large delicate ears it listens for the sound of the insect gnawing within the branch, and is thus able to fix its exact position; with its powerful curved gnawing teeth, as seen in the accompanying figure of the skull, it rapidly cuts away the bark and wood till it exposes the burrow of the insect, most probably the soft larva of some beetle, and then comes into play the extraordinary long
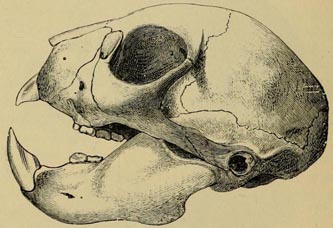
FIG. 42.—AYE-AYE (Chiromys Madagascariensis).
wire-like finger, which enters the small cylindrical burrow, and with the sharp bent claw hooks out the grub. Here we have a most complex adaptation of different parts and organs all converging to one special end, that end being the same as is reached by a group of birds, the woodpeckers, in a different way; and it is a most interesting fact that, although woodpeckers abound in all the great continents, and are especially common in the tropical forests of Asia, Africa, and America, they are quite absent from Madagascar. We may therefore consider that the aye-aye really occupies the same place in nature in the forests of this tropical island, as do the woodpeckers in other parts of the world.
Madagascar also contains the largest and handsomest of the lemurs, consisting of several genera and ten or twelve species, one of the best known being the Ring-tailed lemur (Lemur catta), which is often brought to this

FIG. 43.—RING-TAILED LEMUR (Lemur catta).
country, and is a very delightful pet. It has, however, the habit of springing suddenly on to the shoulders of friends or strangers in a rather alarming manner. A few years ago a specimen (of whose existence near us I was not aware) walked quietly into my garden, and showing
N 2
not the least fear, most unexpectedly jumped upon my shoulder, and thence into a tree under which I was standing. It is about the size of a large cat, but with longer limbs. The woodcut (Fig. 43) gives an excellent idea of this beautiful animal.
There are also several smaller lemurs in Madagascar belonging to the genus Chirogaleus, so that this wonderful island contains twenty-six species of the sub-order, while tropical and South Africa has only sixteen and Asia five. The African species mostly belong to the genus Galago, which consists of small long-tailed active animals, and two others of the genus Perodicticus, which are tailless and confined to the dense forests of equatorial West Africa. All are alike nocturnal in their habits.
Distribution, Affinities, and Zoological Rank of Monkeys.
Having thus sketched an outline of the monkey tribe as regards their more prominent external characters and habits, we must say a few words on their general relations as a distinct order of mammalia. No other group so extensive and so varied as this, is so exclusively tropical in its distribution, a circumstance no doubt due to the fact that monkeys depend so largely on fruit and insects for their subsistence. A very few species extend into the warmer parts of the temperate zones, their extreme limits in the northern hemisphere being Gibraltar, the Western Himalayas at 11,000 feet elevation, East Thibet, and Southern Japan. In America they are found in Mexico, but do not appear to pass beyond the tropic. In the Southern hemisphere they are limited by the extent of the forests in South Brazil, which reach about 30° south latitude. In the East, owing to their entire absence from Australia, they do not reach the southern tropic; but in Africa some baboons range to the southern extremity of the continent.
But this extreme restriction of the order to almost tropical lands is only recent. Directly we go back to the Pliocene period of geology we find the remains of monkeys in France, and even in England. In the earlier
Miocene period several kinds, some of large size, lived in France, Germany, and Greece, all more or less closely allied to living forms of Asia and Africa. About the same period monkeys of the South American type inhabited the United States. In the remote Eocene period the same temperate lands were inhabited by lemurs in the East, and by curious animals believed to be intermediate between lemurs and marmosets in the West. We know from a variety of other evidence that throughout these vast periods a mild and almost sub-tropical climate extended over all Central Europe and parts of North America, while one of a temperate character prevailed as far north as the Arctic circle. The monkey tribe, in these remote periods, enjoyed a far greater range over the earth, and perhaps filled a more important place in Nature than it does now. Its restriction to the comparatively narrow limits of the tropics is no doubt mainly due to the great alteration of climate which occurred at the close of the Tertiary period, but it may have been aided by the continuous development of varied forms of mammalian life better fitted for the contrasted seasons and deciduous vegetation of the north temperate regions. The more extensive area formerly inhabited by the monkey tribe, would have favoured their development into a number of divergent forms in distant regions and adapted to distinct modes of life. As these retreated southward and became concentrated in a more limited area, such as were able to maintain themselves became mingled together as we now find them, the ancient and lowly marmosets and lemurs subsisting side by side with the more recent and more highly developed howlers and anthropoid apes.
Throughout the long ages of the Tertiary period monkeys must have been very abundant and very varied, yet it is but rarely that their fossil remains are found. This, however, is not difficult to explain. The deposits in which mammalian remains most abound are those formed in lakes or in caverns. In the former the bodies of large numbers of terrestrial animals were annually deposited, owing to their having been caught by floods in the tributary streams, swallowed up in marginal bogs or quicksands,
or drowned by the giving way of ice. Caverns were the haunts of hyænas, tigers, bears, and other beasts of prey, which dragged into them the bodies of their victims, and left many of their bones to become imbedded in stalagmite or in the muddy deposit left by floods, while herbivorous animals were often carried into them by these floods, or by falling down the swallow-holes which often open into caverns from above. But, owing to their arboreal habits, monkeys were to a great extent freed from all these dangers. Whether devoured by beasts or birds of prey, or dying a natural death, their bones would usually be left on dry land, where they would slowly decay under atmospheric influences. Only under very exceptional circumstances would they become imbedded in aqueous deposits; and instead of being surprised at their rarity we should rather wonder that so many have been discovered in a fossil state.
Monkeys, as a whole, form a very isolated group, having no near relations to any other mammalia. This is undoubtedly an indication of great antiquity. The peculiar type which has since reached so high a development must have branched off the great mammalian stock at a very remote epoch, certainly as far back as the Secondary period, since in the Eocene we find lemurs and lemurine monkeys already specialised. At this remoter period they were probably not separable from the insectivora, or (perhaps) from the ancestral marsupials. Even now we have one living form, the curious Galeopithecus or flying lemur, which has only recently been separated from the lemurs with which it was formerly united, to be classed as one of the insectivora; and it is only among the Opossums and some other marsupials that we again find hand-like feet with opposable thumbs, which are such a curious and constant feature of the monkey tribe.
This relationship to the lowest of the mammalian tribes seems inconsistent with the place usually accorded to these animals at the head of the entire mammalian series, and opens up the question whether this is a real superiority or whether it depends merely on the obvious relationship to ourselves. If we could suppose a being
gifted with high intelligence, but with a form totally unlike that of man, to have visited the earth before man existed in order to study the various forms of animal life that were found there, we can hardly think he would have placed the monkey tribe so high as we do. He would observe that their whole organisation was specially adapted to an arboreal life, and this specialisation would be rather against their claiming the first rank among terrestrial creatures. Neither in size, nor strength, nor beauty, would they compare with many other forms, while in intelligence they would not surpass, even if they equalled, the horse, the elephant, or the beaver. The carnivora, as a whole, would certainly be held to surpass them in the exquisite perfection of their physical structure, while the flexible trunk of the elephant, combined with his vast strength and admirable sagacity, would probably gain for him the first rank in the animal creation.
But if this would have been a true estimate, the mere fact that the ape is our nearest relation does not necessarily oblige us to come to any other conclusion. Man is undoubtedly the most perfect of all animals, but he is so solely in respect of characters in which he differs from all the monkey tribe—the easily erect posture, the perfect freedom of the hands from all part in locomotion, the large size and complete opposability of the thumb, and the well-developed brain, which enables him fully to utilise these combined physical advantages. The monkeys have none of these; and without them the amount of resemblance they have to us is no advantage, and confers no rank. We are biassed by the too exclusive consideration of the man-like apes. If these did not exist the remaining monkeys could not be thereby deteriorated as to their organisation or lowered in their zoological position, but it is doubtful if we should then class them so high as we now do. We might then dwell more on their resemblances to lower types—to rodents, to insectivora, and to marsupials, and should hardly rank the hideous baboon above the graceful leopard or stately stag. The true conclusion appears to be, that the combination of external characters and internal structure which exists in the monkeys, is that
which, when greatly improved, refined, and beautified, was best calculated to become the perfect instrument of the human intellect and to aid in the development of man's higher nature; while, on the other hand, in the rude, inharmonious, and undeveloped state which it has reached in the quadrumana, it is by no means worthy of the highest place, or can be held to exhibit the most perfect development of sub-human animal life.
CHAPTER VIII
THE DISGUISES OF INSECTS
EVERY one has heard of the wonderful Leaf-insect of the tropics, which is scarcely less a puzzle to the scientific naturalist than it is to the natives of the countries it inhabits. I have been told over and over again by intelligent persons in the East of the curious plant whose leaves changed into insects! And I could never convince them that this was not the true explanation, for they would say, "It is no good your trying to persuade me, for I have seen the creature myself; and I assure you that it has real leaves growing out of it, exactly the same as the other leaves that grow upon the tree." And we really cannot wonder at this belief, for when the creature is alive it remains motionless among the foliage, and the colour, veining, form, and texture of its wing-covers and appendages, are so wonderfully like those of leaves that it is extremely difficult to distinguish it at all.
A few years since a specimen of the Phyllium scythe, the "Walking Leaf" of India, was kept alive at the Royal Botanic Garden at Edinburgh. Mr. Andrew Murray wrote a long account of it, and among other matters says: "It so exactly resembled the leaf on which it fed, that when visitors were shown it, they usually, after looking carefully over the plant for a minute, declared that they could see no insect. It had then to be more minutely pointed out to them; and although seeing is notoriously said to be believing, it looked so absolutely the same as the leaves among which it rested, that this test would rarely satisfy
them; and nothing would convince them that there was a real live insect there but the test of touch—it had to be stirred up to make it move, or taken off the plant to crawl on the finger of the attendant."
But these remarkable insects do not stand alone. There are many others in every order which are "disguised" in a somewhat similar manner, some with equal perfection, others less accurately, but all serving the same purpose—that of protecting the insect from the enemies that would destroy it. I propose now to give a short account of some of the more interesting cases that occur both at home and in the more luxuriant regions of the tropics.
Almost everyone must have noticed the very different way in which the bright colours are distributed in butterflies and moths. In the former, the whole upper surface of the wings is adorned with equally gay colours, while the under surface is always less brilliant, and is generally blotched or mottled with obscure or simple hues. In most moths, on the contrary, the bright colour is restricted to the upper surface of the lower wings, the upper wings being usually of variously mottled brown or ashy tints. This difference is at once seen to be connected with the habits of the insects, the conspicuous colours being so arranged as to be visible during flight, but hidden in repose. On the other hand, the beautiful mottlings and spots and delicate shadings that cause so many moths to resemble bark or lichens, or leaves or twigs, are never developed on those parts of the wings which are hidden during repose. Thus all the Bombycidæ and Noctuidæ, which conceal their hind wings when at rest, have them either quite plain or ornamented with rich orange or crimson hues; while almost all the Geometridæ, which rest with their wings spread open, and such of the Bombycidæ as have the same habit, are mottled and tinted alike on both front and hind wings.
These general facts as to the distribution of colour constitute the first stage in that process of "disguise" which becomes so wonderfully developed in a few conspicuous cases. The next stage is exhibited by the fact that there is a general agreement between the colour of a large
number of moths and the prevailing tints of nature at the season when they appear. Out of fifty-two autumn-flying moths, it has been noticed that a large proportion are of various tints of yellow and brown, so as exactly to match with the "sere and yellow leaf;" while in winter they are of grey and silvery tints, like the washed-out leaves and grass, the fog and the hoar-frost, which give a tone to every landscape at this season.
We now come to a closer and more special disguise. Many of the moths that rest during the day on palings or on the trunks of trees are marked and coloured so as to
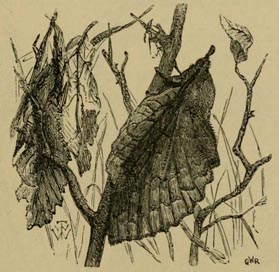
FIG 44.—LAPPET MOTH.
match the tints of the bark and lichens, and thus to escape observation. As examples of this numerous class, we may mention two of our commonest species—the "Dagger" (Acronycta psi) and the pretty green Agriopis aprilina. The Lappet moth (Gastropacha quercifolia) when at rest resembles very closely a small bunch of dead leaves; and at a little distance could hardly be taken to be a moth, so curiously does it spread out its hind wings so as to project beyond the others. The accompanying cut (Fig. 44) by the late Mr. T. W. Wood, is an accurate representation of this insect in its attitude of repose.
One of the most curious of these resemblances is that of the Buff-tip moth (Pygæra bucephala). This insect closes its wings so as almost to form a cylinder; and on the tip of each wing is an oval yellowish spot, edged with a dark brown double line. The wings are greyish and hoary; and the head, again, is much contracted beneath the large thorax, which is also of a buff colour, with a double brown marginal line. The result of this arrangement is that the insect looks at first sight like a piece of

FIG. 45.—BUFF-TIP MOTH.
stick, one end being broken off nearly square, the other end more obliquely (Fig. 45); and as it often rests on the ground, among grass or on leaves, it may easily be mistaken for a piece of a broken branch which has fallen to the ground. Many more of these beautiful adaptations remain to be discovered in our native insects. That most elegant insect, the Elephant Hawk moth, is of a reddish-pink colour, mingled with dull yellowish-green, and with specks and streaks of white; but it has not been noticed
how closely all these colours must assimilate it to the handsome red-flowered Willow-herb (Epilobium), on which the larva feeds, and on which the female insect, while depositing her eggs, no doubt often reposes. The petals of the common Epilobium angustifolium, for instance, are of the same pinky-red as the moth; its stems and seed-pods are green, tinted with brown-purple or yellowish, while the white filaments of its stamens correspond to the white lines and streaks of the insect. It is evident, therefore, that while reposing amid a clump of these plants, the Elephant Hawk-moth, although so brilliantly coloured, must be exceedingly difficult to detect, since every part of its body is of exactly the same hue as some portion of the flowers.
We owe the discovery of one of the most beautiful examples of "disguise" in a native insect to the talented young artist and close observer of nature who furnished the illustrations for this chapter. He tells us that one fine afternoon in May, being overtaken by a shower, he sought shelter under a hedge, where, among other flowers, the wild parsley (Anthriscus sylvestris) grew in the greatest profusion. While observing the light and elegant forms of these plants, he noticed what appeared to be a small bunch of flowers projecting beyond the rest; and a closer examination led him to the interesting discovery that our beautiful little "Orange-tip," one of the gayest and brightest of our native butterflies, was reposing among these flowers in such a manner as to gain a complete protection by its resemblance to them. He was now able to see the use of almost every detail, both of the form and colouring of this insect. The bright orange patch, so beautiful when the insect is on the wing, was hidden beneath the hind wings; and there is thus a very good reason for the circumstance that the colour does not extend so far on the under as on the upper surface of the wing, and also for the tip of the upper wings being mottled with green beneath, like the whole surface of the hind wings; for as they are a little longer than the hind wings, the whole form one uniformly coloured surface when the
wings are closed (Fig. 46). Various other species of Anthocharis, as well as the pretty Zegris of Eastern Europe and our rare Pieris daplidice, are coloured in a

FIG. 46.—ORANGE-TIP BUTTERFLY.
similar manner on the under side, though with varying degrees of brilliancy; and it is probable that they are accustomed to repose on the flowers of umbelliferous or cruciferous plants of suitable colours. One of the hand-
somest species of Indian Pieridæ, the Iphias glaucippe, whose upper wings are ornamented above with a large patch of vivid orange-red, is so tinted and mottled on the under side as to resemble a dry leaf; and a strong dark line running through the centre of the wings represents the midrib. This species often rests on the ground, on the banks of streams or on beds of gravel, and depresses the upper wings so much between the lower ones as to form an outline very similar to that of a leaf; and this is no doubt a great protection to it; for although so large and showy an insect, it is very plentiful.
By far the most singular and most perfect disguise I have ever met with in a Lepidopterous insect is that of a common Indian butterfly, Kallima inachis, and its Malayan ally Kallima paralekta. I had the satisfaction of observing the habits of the latter in Sumatra, where it is rather plentiful at the end of the dry season. It is a large and showy insect when on the wing, the upper surface being glossed with blue and purple, and the fore wings crossed obliquely by a broad band of rich orange. The under surface of the wings is totally different, and is seen at a glance to resemble a dead leaf. The hind wings terminate in a little tail, which forms the stalk of the leaf, and from this to the apex is a slightly curved dark brown line representing the midrib. The transverse striæ which cross the discoidal cell in many butterflies are here continued so as to form lateral veins, and the usual sub-marginal striæ on the hind wings, slightly modified, represent others towards the base of the wing. But it is only when the habits of the insect are observed that the disguise becomes manifested in all its perfection (Fig. 47). This butterfly, like many others, has the habit of resting only upon a nearly vertical twig or branch, with the wings closed together so as completely to conceal the upper surface. In this position, the little tail of the hind wings exactly touches the branch, and we now see why it is always curved inwards a little; for if it were quite straight, it would hang clear of the branch, and thus fail to represent an attached leaf. There is a little scallop or hollow on the margin of the fore wings at the base, which serves
to conceal the head of the butterfly, which is very small for its size, and the long antennæ are carried back and

FIG. 47.—LEAF BUTTERFLIES AT REST AND FLYING.
hidden between the folded wings. When sitting on a twig in the manner described, the insect is to all appearance a perfect dry leaf,—yet it is evident that its chances of
escape would be much increased if it were surrounded by real dry leaves instead of by green ones; for if, when pursued, it took shelter in a growing bush, it could hardly fail to be still a conspicuous object. Marvellous to relate, it does possess the habit of almost invariably entering a bush loaded with dead leaves, and is so instantly lost to sight, owing to its close resemblance to all the surrounding objects, that I doubt if the most vigilant fly-catcher could detect it. I have myself often been utterly puzzled. I have watched it settle, apparently in a very conspicuous situation, a few yards off, but on crawling carefully up to the spot have been quite unable to detect any living thing. Sometimes, while gazing intently, a butterfly would start out from just before my eyes, and again enter another dead bush a few yards off, again to be lost in the same manner. Once or twice only was I able to detect it sitting, and admire the wonderful disguise which a most strange combination of colour, form, and habits enabled it instantaneously to assume. But there is yet another peculiarity which adds to the concealment of this species. Scarcely two of the specimens are alike in colour on the under side, but vary through all the shades of pale buff, yellow, brown, and deep rusty orange which dried leaves assume. Others are speckled over with little black dots like mildewed leaves, or have clusters of spots or irregular blotches, like the minute fungi that attack dead leaves; so that a dozen of these insects might settle on a perfectly bare spray, and clothe it at once with withered foliage not distinguishable from that of the surrounding branches!
The protection derived from a vegetable disguise is not confined to the perfect Lepidoptera, but is often equally remarkable in their larvæ. The caterpillar of a European moth that feeds on the privet (Hadena ligustri) is so exactly the colour of the under side of the leaf, on which it sits in the day-time, that you may have the leaf in your hand and yet not discover it. In the caterpillars of the Geometridæ, form, colour, and habit combine to disguise many of the species. Those of the Brimstone and Swallowtail moths may be taken as examples. They have the
VOL. I. O
habit of stretching themselves out obliquely when in repose, attached only by the clasping legs at the further extremity, and will remain stiff and motionless in this position for hours. The little protuberances on the body, their colour and attitude, give them so exactly the appearance of twigs of the living tree, that we may easily conceive the advantage this disguise must be to them; for it is certain that many will escape destruction when more conspicuous insects will be devoured.
Among the extensive group of the Coleoptera, the examples of a protective disguise are literally innumerable. In the tropics, every fallen tree swarms with beetles, and a large number of these so closely resemble the bark to which they cling, that it requires a close examination to detect them. The families of the Longicorns and Curculios furnish the greater part of these; and among the former, that which wears perhaps the most perfect disguise is the Onychocerus scorpio. This beetle is common in South America, and was found abundantly by Mr. Bates on the banks of the Amazon, but always clinging to the rough bark of one kind of tree, called by the natives Tapiribà. This bark was so closely imitated by the beetle itself,—its elytra and thorax being tubercled and coloured so as exactly to match it, and the insect clinging so closely as to form, apparently, one surface with the tree, that Mr. Bates assures me it was often absolutely impossible to detect it by the closest inspection as long as it remained motionless!
Many of the Tiger beetles, although they are such conspicuous and beautiful objects in our cabinets, are well disguised when in their natural stations. Our commonest species, Cicindela campestris, is fond of grassy banks, where its green colour makes it difficult to see it. Cicindela maritima is almost exactly the same colour as the sandy shores it haunts. The large Cicindela heros frequents the mountainous forests of Celebes, where its brown colour exactly matches with the dead leaves that cover the ground. The magnificent velvety-green Cicindela gloriosa was captured only on wet moss-covered rocks in the bed of a mountain torrent in the island of Celebes, where it
was very difficult to see it. The pale-coloured Cicindela Durvillei was found on coral sand of almost exactly its own colour; and I noticed generally that, whatever the colour of the sand or the soil, the common Tiger beetles of the locality were of the same hue. A most remarkable instance of this was a species which I found only on the glistening, slimy mud of salt marshes, the colour and shine of which it matched so exactly that at a few yards' distance I could only detect it by the shadow it cast when the sun shone!
Several Buprestidæ of the genus Corœbus resemble the dung of birds freshly dropped on leaves, and I have often been puzzled to determine whether what I saw was worth picking up or not. Mr. Bates tells us that Chlamys pilula cannot be distinguished from the dung of caterpillars. Our own Onthophilus sulcatus is very like the seed of an umbelliferous plant, and the common Pill beetle (Byrrhus pilula) would be taken for anything rather than an insect.
We must now turn to the Orthopterous insects, which contain some of the most surprising cases of disguise yet discovered. The true Walking Leaf has been already described at the commencement of this chapter, but there are other insects of a quite different structure which almost equally resemble leaves, as shown by the names given to them by the old writers; such as Locusta citrifolia, L. laurifolia, L. myrtifolia, &c. Acrydium gallinaceum, from the Malay Archipelago, has an immense erect leaf-like thorax; A. platypterum has wings like the most beautiful smooth green leaves; while A. gibbosum is like a little shapeless lump of mud or stick. The voracious Mantidæ are often concealed in a similar manner. Many have the thorax broadly dilated, and, with the wing-covers, coloured like a dead or a green leaf; and one has large brown legs and small wings, so that it looks more like a cluster of bits of stick and withered leaves than a living insect.
The true Phasmidæ, or Stick-insects, are the most curious, perhaps, of all, and they are much more abundant in the eastern forests than the Leaf-insects. They vary from a few inches to a foot long, and are almost always of the colour and shape of pieces of stick, the legs forming
O 2
FIG 48.—STICK INSECT.
the branches. One of the most curious facts connected with them is that they seem to know that if they rested in the symmetrical attitudes in which they are always drawn, with their legs spread out uniformly on each side, they would soon be detected. They are accordingly found stretched out motionless in the most unsymmetrical manner possible—one leg out on one side, and two on the other, for example, the remaining legs fitting so closely to the body that they appear to form one piece with it.
They lay generally across leaves and twigs, as if they had accidentally fallen there from some dry branch overhead; and so impossible is it to detect them by the eye that I used to make it a practice, when walking along in the forests, to touch every suspicious bit of dead stick I saw loose on the foliage, as the only means of finding out whether they were real sticks or Stick-insects. Sometimes they are exactly the colour of lichen-covered branches, and are covered with little foliaceous expansions. One that inhabits the swampy forests of Borneo has these of a beautiful olive-green colour, so as exactly to resemble a creeping moss or jungermannia; and the Dyak who brought it me assured me it was very curious, for he had never before seen an insect grown all over with moss while alive! I was quite as much astonished as he was, for I could hardly believe my eyes, and it was only after close and repeated examination that I could convince myself it was not a real plant that covered the animal. This insect loses all its beauty when dried, and it has been very poorly figured by the Dutch naturalists, and very inappropriately named Ceroxylus laceratus, from its torn and shaggy appearance in the preserved specimens.
In the deserts of Egypt are some curious Mantidæ which are so exactly the colour of the soil they live upon that the closest inspection can scarcely detect them. It is even stated that where the soil changes from brown to white or yellow in a few yards' distance, the insects change also, and always correspond in colour to their habitation. The caterpillar of a European moth, Bryophila algæ, is said to change in a similar manner, being yellow when found on the yellow Lichen juniperinus, but grey when on the grey Lichen saxatilis. In this case, however, the food may probably produce the change of colour, as it is known to do in some other larvæ. Some cases more to the point have been observed by the late Mr. T. W. Wood. He states that the chrysalis of the common Tortoise-shell butterfly is of a very different colour according to its position. When attached to a nettle, it is of a golden colour; when on a wall or fence, mottled grey; and when on a tarred paling, nearly black. Once he placed some
larvæ of the Swallow-tailed butterfly in chip boxes, where they changed into chrysalids; but, strange to say, instead of being green or dusky, as they usually are, they were of exactly the same colour as the inside of the box, without any marking whatever. Some of them produced very fine butterflies, which shows that they were healthy.1
These curious facts prove that we have yet much to learn as to the causes which determine the colours of animals, and it is to be wished that a few of Our young naturalists would experiment on some of our commonest insects, rearing them from the egg exposed to the influence of differently coloured objects and carefully registering the result. In this chapter I have only desired to call attention to some curious facts in the colouring of insects, and more especially to the disguises which serve to protect them from their enemies, or enable them more easily to entrap their prey. Such of my readers as may wish to know more of this subject, and may desire to learn how these strange modifications of form and colour have probably arisen, are referred to Chapter III. of my Natural Selection and Tropical Nature, in which the most recent views of Mr. Darwin's disciples are fully explained.
1 Since the present chapter was written this subject has been well investigated by Prof. E. B. Poulton and other entomologists. See his book on The Colours of Animals, Chaps. VIII. and IX.
CHAPTER IX
ENGLISH AND AMERICAN FLOWERS
THE numerous English writers who have described their impressions of North America tell us all about the people, their manners and customs, their hotels and churches, the mode of travelling and the scenery, the rivers and waterfalls, the mountains and forests, the prairies and deserts; but hardly ever do they give us any information as to the kind of vegetation that covers the surface of the soil, or the flowers that adorn the roadsides, the forests, or the mountains. Hence it comes to pass that the majority of English readers, even those who delight in the wild flowers of their own country or the more varied beauties of the Alpine flora, have usually the vaguest and most erroneous ideas as to what flowers are to be found in the United States and Canada, and to what extent they resemble or differ from those of our own country.
There are many circumstances which render it difficult, even for the native who is not a botanist, to learn much about American wild flowers. Confining ourselves at present to the North-Eastern States, we may say that three hundred years ago the whole country was covered with forest, and, with few exceptions, the herbaceous flowering plants were such as grew in the shade of trees or in the few open glades, in bogs, or on the banks of streams. Now, these forests have been so completely cleared away that comparatively little remains in its primitive condition, and often over extensive areas hardly a
patch of original woodland is to be found. In other districts there is plenty of land covered with trees, but these are usually new growths, the timber having been felled again and again, as required for firewood, for fencing, or for other purposes. This wholesale clearing of the original forest-covering of the soil has led, no doubt, to the destruction of many lowly plants, some of which have become exterminated altogether, while others have been able to survive only in the few spots that still offer suitable conditions for their existence. Such places are comparatively rare, and often difficult of access; and hence the country, for a considerable distance round the larger cities and towns, affords but few of the really native plants, while common European weeds often abound. The old hedgerows, the shady banks and moist ditches, the deep-cut lanes, and the numerous footpaths of our own country, which afford abundant stations where wild flowers have been preserved to us from prehistoric times, are almost wholly wanting in America. There the seeker after wild flowers must usually be prepared to walk long distances over rough and pathless fields or hills in order to reach the places where alone he has any chance of finding the rarer or the more beautiful species. Owing to this absence of pleasant rural pathways the inhabitants of the towns rarely walk far into the country for exercise or pleasure unless they have some special pursuit of sport or natural history, and that want of interest in the natural productions of the district which is sufficiently common in England is still more prevalent in America.
The relations of the entire flora of temperate North America to that of Europe and Northern Asia have been the subject of much discussion among botanists. The late Professor Asa Gray made known, and, to some extent, popularised, the curious anomalies which these relations present, especially as regards the close affinity of the plants (more especially of the trees and shrubs) of the Eastern United States with those of Eastern Asia and Japan. Some account of Asa Gray's researches was given by myself, in 1878, in an article on "Epping Forest" (which is reprinted in Vol. II. of this work), and they are only
now referred to because they have been used to uphold the theory that, as regards the distribution of plants, the north temperate zone of the eastern and western hemispheres cannot be separated, but must be considered to form one botanical region. Recently, however, Sir Joseph Hooker has stated his opinion that if we go beyond the two fundamental botanical divisions—the tropical and temperate floras, which, for purposes of geographical distribution, are of little interest, we must consider that the temperate floras of the Old and New Worlds are as distinct as are the tropical floras of the same areas; and he adds that, although the resemblances as regards certain genera and species of plants between Eastern America and Eastern Asia, is very remarkable, yet the temperate floras of Asia and America are in other respects totally dissimilar.1 In the present chapter I shall endeavour to show, in a popular manner, what is the nature and extent of the dissimilarity between America and Europe as regards what are commonly known as wild flowers.
The Flowers of Europe, and the Eastern States Compared.
In order to restrict the inquiry within moderate limits, it is proposed to consider, first and mainly, the relations between the wild flowers of Great Britain as representing those of Western Europe, and those of Eastern North America as given in Asa Gray's Botany of the Northern United States, which includes the country from New England to Wisconsin, and southward to Ohio and Pennsylvania. This area may be fairly compared with that of England, France, and Germany, and will serve as a foundation for the wider comparison between North America east of the Mississippi with Europe, or of the whole of temperate North America with temperate Europe and Asia, to which occasional reference will have to be made. It must be understood that as our comparison regards only the native plants of the two countries, those numerous British or European species which have been introduced into America by human agency and have often
1 See Biologia Centrali-Americana, Botany, vol. i., pp. lxvi—lxvii.
become common weeds, will be left out of consideration altogether. We have to do only with the condition of the vegetation brought about by nature, undisturbed by the effects which have recently been produced by man.
There are two separate phenomena by which we may estimate the relations of the floras of two countries, both of which are important factors in the comparison—the absence from one country of whole groups of plants which are both common and widespread in the other, and the presence of new types entirely unknown in the other. It is usual to lay much more stress on the latter phenomenon, because the former occurs when there is no essential difference between the floras, the one having been recently derived from the other. Thus, many species, and even genera, of West European plants are absent from Britain, but this does not lead us to consider the British flora as being essentially different from that of Europe, the deficiencies being plainly attributable to the smaller area, the limited range of climate, the recent glacial epoch, and other such causes. But, when the country in which the deficiency occurs is fairly comparable with the other in all these respects, the cause of the phenomenon is evidently a deep-seated one, and must be held to show a fundamental diversity in their floras.
There are, of course, in every extensive flora such as that of North America a considerable number of almost cosmopolitan groups or species, and many others which are found in all temperate regions. Thus, no less than 115 European genera and 58 European species are found at the antipodes in New Zealand, and many others in Australia and South temperate America. Among these are such familiar plants as buttercups, anemones, poppies, violets, St. John's worts, gentians, forget-me-nots, many genera of cresses and other crucifers, mint, skull-cap, loose-strife, sea-lavender, and many others; and there are also in the same remote countries such common English species as the lady's-smock (Cardamine pratensis), chickweed (Stellaria media), the cut-leaved geranium (Geranium dissectum), the silver-weed (Potentilla anserina), the common bind-weed (Calystegia sepium), and scores of others, all considered to be
indigenous and not introduced by man; yet the differences between the European and Australian floras are very great, and are hardly surpassed by those of any two regions on the globe. It is evident, therefore, that we must expect to find a considerable number of English species in North America, and a still larger number of English genera, because this is a feature which occurs in all temperate regions, and cannot be held to prove any special relationship between these two countries. Among these familiar English flowers we find a tolerable number of violets, anemones, St. John's worts, vetches, potentillas, willow-herbs, gentians, and some others; while wild geraniums, saxifrages, stonecrops, campanulas, forget-me-nots, and true orchises are far less frequently met with than with us.
But what most strikes the English botanist (next to the altogether unfamiliar types that everywhere abound) is the total absence or extreme rarity of many plants and groups of plants which are the most abundant and familiar of our native flowers, and which are almost equally common throughout Europe, and often throughout northern Asia. There are, for instance, no true poppies like those so abundant in our corn-fields; no common or musk-mallows of the genus Malva, or gorse or broom or rest-harrow; no teasel or scabious; no true heaths; no bugloss or comfrey; no ivy to adorn the old trees and walls with its glossy foliage; no mullein, toad-flax,1 snapdragon, or fox-glove; no scented thyme, basil or marjoram; no bright blue ground-ivy or bugle; no white or yellow or purple dead-nettles; no scarlet pimpernel; not even a primrose or a cowslip in all the land. There are, it is true, two species of Primula in the North-Eastern States, one the bird's-eye primrose of our northern counties, and another still smaller peculiar species, but both are confined to limited districts near the great lakes, and are not to be found unless specially searched for; and no other primroses are to be met with till we reach the Rocky Mountains, where there are two or three high alpine species.
1 Toad-flax (Lenaria vulgaris) is common in some parts of New England, but it is an introduced species run wild.
Coming now to the endogenous plants, we find equally remarkable deficiencies. No daffodil, snowdrop, or snow-flake is to be found wild in all North America, neither is there any crocus, wild hyacinth, colchicum, or lily-of-the-valley. The beautiful genus Ophrys, containing our bee, fly, and spider orchises, is quite unknown; and such familiar plants as the black-briony of our hedges, the flowering-rush of our streams and ditches, and the curious butcher's broom of our dry woods, are nowhere to be met with.
Now the important thing to be noted is, that most of these plants are not only abundant and familiar in many parts of England, but are widely spread throughout Europe, and the larger part of them belong to groups which extend into Northern Asia, and often reach the eastern extremity of that continent. If we were to include less important or less familiar plants this list might be doubled or trebled; and it might be still further extended if we took account of genera which range widely over Europe and Asia, but happen to be rare or altogether wanting in England. Such, for example, are the following well-known garden flowers. The white and yellow asphodels of South Europe; the red valerian (Centranthus), naturalised in many places on our chalk cliffs and old walls; the cinerarias, the gum-cistuses, the cyclamens, and the daphnes; the true pinks (Dianthus); the numerous dwarf brooms (Genista); the corn-flags (Gladiolus); the candytufts (Iberis); the lavender and the rosemary; the ox-eye daisies; the stocks; the Star-of-Bethlehem; the pœonies; the mignonettes; the garden rue; the various soap-worts; the tulips, the periwinkles, and a hundred others.
It must always be remembered, that the British plants noticed above as being absent from the indigenous flora of the United States are abundant with us and form characteristic features of our flora; that the larger portion of them range widely over Europe and Western Asia; that more than half of them extend across Northern Asia to the Pacific and often to China and Japan; while several extend over the greater portion of the eastern hemi-
sphere, and are found also in Australia or South Africa. The absence of such a number of the characteristic and dominant groups of plants of the temperate zone from so extensive and varied an area as the Eastern United States and Canada, is of itself a very remarkable phenomenon, and affords a primâ facie ground for treating the temperate regions of the New World as a distinct botanical region.
Another feature to which botanists attach much importance in the comparison of separate floras is the proportionate abundance of the various orders and tribes in the two countries, which, when very different, leads to the general vegetation having a distinctive aspect. In this respect, Europe and Eastern America differ greatly. Among the most abundant and characteristic groups, which every one recognises in our own country and in Europe as common plants everywhere to be met with, are those of the cabbage and cress tribe (Cruciferæ), the pink family (Caryophyllaceæ), the umbel-bearers (Umbel-liferæ), the thistle-tribe of the Compositæ, the bluebells (Campanulaceæ), the primroses (Primulaceæ), and the orchises (Orchidacæ); but all these are much less frequent in North America, and are usually so scarce as to take little or no part in determining the special aspect of the vegetation. As an illustration of this difference, there are only twelve indigenous genera of Cruciferæ in the North-Eastern United States with about thirty-five indigenous species, while the comparatively poor British flora possesses twenty-four genera, and fifty-four species.
Instead of these characteristic European types we have in America some peculiar Rubiaceæ, among which is the pretty creeping Mitchella or partridge berry; and an enormous preponderance of Compositæ, including numbers of non-European genera and a great variety of eupatoriums, asters, golden-rods, and sun-flowers, together with some of our well-known garden flowers such as Liatris, Rudbeckia and Coreopsis. The milk-worts (Polygala), are rather numerous, and the milk-weeds (Asclepias) still more so, and these last are quite unlike any European plants. The beautiful phloxes are a very characteristic type almost exclusively confined to North America, and
often affording brilliant patches of floral colours. Among the endogenous plants the numerous specie of Smilax, well called "green-briar," are highly characteristic and peculiar, though the genus is found in Southern Europe, while the beautiful wood-lilies of the genus Trillium (found again in Eastern Asia) have curious or ornamental flowers. Add to these the interesting spider-worts forming the genera Commelyna and Tradescantia, and having their allies in the tropics, and we have completed the enumeration of the more conspicuous groups of non-European herbaceous flowering plants which give a distinctive character to the flora.
There are, however, many other plants, which, although belonging to small orders and not represented by more than one or a very few species, are yet so abundant in individuals, and so widely spread over the country, as to contribute largely to the general impression which the North American flora produces on an English botanist on his first visit to the country. This may be illustrated by a brief account of some of the present writer's rambles in search of American flowers.
Rambles after American Flowers.
My first walk was on the 13th February, in the woody country to the north-west of Washington under the guidance of my friend, Professor Lester Ward. Here we found on dry banks the beautiful little May-flower (Epigæa repens), and the pretty spotted wintergreen (Chimaphila maculata), both members of the heath family, and both of genera almost peculiar to America, allied species of each being found in Japan; while some of the forest slopes were covered with the beautiful Kalmia latifolia, another peculiarly American genus of Ericaceæ. The curious parasitic "beech-drops," one of the Orobanche tribe, is also peculiar, while the "partridge-berry" (Mitchella repens) has its only ally in Japan. Other plants observed were the Aralia spinosa or Hercules' Club, a small tree of a non-European genus; a species of Smilax like a slender leafless rose-bush; a hairy-leaved blackberry (Rubus villosus); a wild vine; a white hepatica in flower identical
with the European species; the curious skunk-cabbage (Symplocarpus fætidus), belonging to the Arum family and also in flower; the "blazing-star" (Chamælirium luteum), one of the colchicum tribe and peculiar to North America, the rattle-snake plantain (Goodyera pubescens), really an orchis, of which there is one European species found also in Scotland, and a handsome sedge (Carex platyphylla). Thus, out of fourteen plants distinguished at this wintry season, only three belonged to British, and four or five to European genera, while the large majority were either quite peculiar to America or only found elsewhere in Japan, Eastern Asia, or the Pacific Islands. During another excursion to the rich locality of High Island, five miles from Washington, on March 27th, several other interesting and characteristic plants were found. Such were the "spring beauty" (Claytonia virginica), a pretty little rosy white flower belonging to the Portulaceæ which often carpets the woods and takes the place of our wood-anemone, for though there are several wood-anemones in America they do not form so important a feature of the spring vegetation as with us. The rare and pretty "harbinger of spring" (Erigenia bulbosa), a minute umbelliferous plant quite peculiar to America, grew here abundantly, as did the pinnate-leaved Virginian water-leaf (Hydrophyllum virginianum). The may-apple (Podophyllum peltatum), and the twin-leaf (Jeffersonia diphylla), herbaceous plants of the Berberis family, were abundant, the former occurring elsewhere only in the Himalayas, while the latter is North American and East Asiatic. A yellow violet, a perennial chickweed, a tooth-wort (Dentaria), a stone-crop, and an inconspicuous saxifrage, alone showed any resemblance to our native vegetation.
About the middle of April, in the vicinity of Cincinnati, I was introduced to the spring flowers of the North-Eastern States, in their full development. The woods were here carpeted in places with the "spring beauty," while in other parts there were sheets of the curious "Dutchman's breeches" (Dicentra cucullaria), like a small yellow dielytra, to which it is allied. Then there were smaller patches of the Thalictrum anemonoides, resembling a very
slender wood-anemone; the exquisite little "Blue-eyed Mary," sometimes called "Innocence" (Collinsia verna); the handsome celandine-poppy (Stylophorum diphyllum), like our "greater celandine," but with larger and more richly coloured flowers; the elegant lilac-coloured Phlox divaricata; and the "blood-root" (Sanguinaria canadensis), with its beautiful white star-shaped flowers.
Here, too, the buds of the handsome purple wood-lily (Trillium erectum) were just showing themselves, and there were large patches of the yellow and white American dog's-tooth violet (Erythronium Americanum), just coming into bloom. In a damp river-bottom, the exquisite blue Mertensia virginiea was found. It is called here the "Virginian cowslip," its drooping porcelain-blue bells being somewhat of the size and form of those of the true cowslip, but the plant is really allied to our lungworts. More homely-looking plants were a creeping yellow buttercup, with blue, white, and yellow-flowered violets, but they were utterly insignificant as compared with the many new and strange forms that constituted the bulk of the vegetation.
At the end of July I had the opportunity of seeing the swampy forests of Michigan, with their abundance of ferns, their pitcher plants (Sarracenia), yellow-fringed orchises (Habenaria ciliaris), and the curious little gold-thread (Coptis trifolia), found also in Arctic Europe, and so named from its yellow thread-like roots,—all three growing in the dense carpet of sphagnum moss which covers the ground to the depth of one or two feet. In the cleared marshy ground, and along the margins of the streams and ditches, was a dense vegetation of asters, golden-rods, and other composites, many of which were of genera unknown in Britain or in Europe, while still lingering on the burnt-up road sides were the handsome flowering spurge (Euphorbia corollata), with its curious white flowers, and the elegant foliage of the bird's-foot violet.
General Aspect of American Rural Scenery.
A few remarks on the general aspects of the country as regards native vegetation and flowers must conclude this very imperfect sketch. What most impresses the nature-loving Englishman while travelling in America is, the newness and rawness of the country, and the almost universal absence of that harmonious interblending of wild nature with human cultivation, which is so charming over a large part of England. In these North-Eastern States, the native forests have been so ruthlessly destroyed that fine trees are comparatively rare, and such noble elms, beeches, oaks, and sycamores as are to be found arching over the lanes and shading the farmhouses and cottages in a thousand English villages, are only to be seen near a few of the towns in the older settled States, or as isolated specimens which are regarded as something remarkable. Instead of the old hedgerows with tall elms, spreading oaks, and an occasional beech, hornbeam, birch, or holly, we see everywhere the ugly snake-fence of split rails, or the still more unsightly boundary of barbed wire. Owing to the country being mostly cut up into one-mile square sections, subdivided into quarters, along the outer boundaries of which is the only right-of-way for access to the different farms, the chief country roads or tracks zigzag along these section-lines without any regard to the contours of the land. It is probably owing to the cost of labour and the necessity of bringing large areas under cultivation as quickly as possible, that our system of fencing by live hedges, growing on a bank, with a ditch on one side for drainage, seems to be absolutely unknown in America; and hence the constant references of English writers on rural scenery and customs to "the ditch," or "the hedge," are unintelligible to most Americans.
The extreme rapidity with which the land has been cleared of its original forest seems to have favoured the spread of imported weeds, many of which are specially adapted to seize upon and monopolise newly exposed or loosened soil; and this has prevented the native plants,
VOL. I. P
which might have adapted themselves to the new conditions had the change gone on very slowly, from gaining a footing. Hence it is that the cultivated fields and the artificial grass lands are less flowery than our hedge-bordered fields and old pastures, while the railway banks never exhibit such displays of floral beauty as they often do with us. An American writer in The Century for June, 1887, summarises the general result of these varied causes, with a severe truthfulness that would hardly be courteous in a stranger, in the following words:—
"A whole huge continent has been so touched by human hands, that over a large part of its surface it has been reduced to a state of unkempt, sordid ugliness; and it can be brought back into a state of beauty only by further touches of the same hands more intelligently applied."
Let us hope that intelligence of this kind will soon be cultivated as an essential part of education in all American schools. This alone will, however, have no effect so long as the fierce competition of great capitalists, farmers and manufacturers, reduces the actual cultivator of the soil, whether owner, tenant, or labourer, to a condition of sordid poverty, and a life of grinding labour which leaves neither leisure nor desire for the creation or preservation of natural beauty in his surroundings.1
Floral Beauty in England and America.
Although with the limited opportunities afforded by one spring and summer spent in America, it is impossible to speak with certainty, yet both from my own observation, and from information received from residents in various parts of the Eastern States, it seems to me, that in no part of America, east of the Mississippi, is there such a succession of floral beauty and display of exquisite colour as are to be found in many parts of England. Such, for instance, are the woods and fields of daffodils, "which come
1 American periodicals are full of accounts and illustrations of the poverty and hard lives of the small farmers. See, in The Arena of July, the article by Hamlin Garland, A Prairie Heroine, and the same writer's volume, Main Travelled Roads.
before the swallow dares, and take the winds of March with beauty"; the wild hyacinths, whose nodding bells, of exquisite form and colour individually, carpet our woods in April with sheets of the purest azure; the soft yellow of primroses in coppices or along sunny hedge banks; the rich golden yellow of the gorse-bushes which, when seen in perfection as in the Isle of Wight, Cornwall, or Ireland, is so superlatively glorious, that we cannot wonder at the enthusiasm of the great Linnæus, who, on beholding it, knelt down and thanked God for so much beauty; later on, the clearer yellow of the bloom is hardly less brilliant on our heaths and railway banks, while the red ragged-robin, and the purple or rosy orchises often adorn our marshes and meadows with masses of colour; then come the fields and dry slopes, gay with scarlet poppies, and the noble spikes of foxgloves in the copses and on rough banks, followed by, perhaps, the most exquisitely beautiful sight of all, the brilliant sheets and patches of purple heath, sometimes alternating with the tender green of the young bracken, as on some of the mountain slopes in Wales, sometimes intermingled with the rich golden clumps of the dwarf gorse, as on the wild heaths of Surrey or Dorset.
Truly, the Englishman has no need to go abroad to revel in the beauty of colour as produced by flowers. Although the number of species of plants which inhabit our islands is far less than in most continental areas of equal extent, although the gloom and grey of our skies is proverbial and we miss the bright sunshine of American or Eastern summers, yet these deficiencies do not appear to lessen the luxuriant display of bright colours in our native plants. The mountains of Switzerland, the arid plains of the Cape and of Australia, the forests and swamps of North America, provide us with thousands of beautiful flowers for the adornment of our gardens and greenhouses, yet, from the descriptions of these countries by travellers or by residents, it does not seem that any one of them produces a succession of floral pictures to surpass, or even to equal, those which the changing seasons display before us at our very doors. The absence of fierce, long-continued
P 2
sunshine, which renders it difficult for us to grow many fruits and flowers which flourish even in the short Canadian summer, lengthens out our seasons favourable to vegetation, so that from the violets and daffodils of March, to the heaths and campanulas, the knapweeds, and the scabious of September or October, we are never without some added charm to our country walks if we choose to search out the appropriate spots where the flowers of each month add their bright colours to the landscape.
To the botanist, the poverty of our English flora contrasts unfavourably with the number of species and the strange or beautiful forms to be found in many other temperate regions, and to him it is a great delight to make the acquaintance, for the first time, in their native wilds, of the many curious plants which he has only known before in gardens or in herbaria. But the simple lover of flowers, both for their individual beauty and for the charm of colour they add to the landscape, may rest assured that, perhaps with the single exception of Switzerland, few temperate countries can equal, while none can very much surpass his own.
CHAPTER X
FLOWERS AND FORESTS OF THE FAR WEST
TEMPERATE North America, as regards its types of vegetation, consists of four well-marked subdivisions. The most important and the richest in species is the great forest region of the Eastern States, whose main peculiarities were indicated in the preceding chapter. West of this area, and extending from a short distance beyond the Mississippi to the base of the Rocky Mountains is the region of the great plains, almost destitute of trees, except in the river bottoms, but with a fairly rich herbaceous flora; and a very similar vegetation is found in the half-desert valleys and plains between the Rocky Mountains and the Sierra Nevada. A third botanical district consists of the higher wooded portions of the Rocky Mountains, together with the peaks and high valleys above the timber-line, in which the vegetation is, in many respects, very distinct from that of any other part of temperate America. Lastly comes the Californian region, extending from the Pacific coast to the upper limit of trees in the Sierra Nevada, a country of surpassing interest to the botanist, and well-known to every lover of flowers for the great number of beautiful and peculiar forms it has furnished to our gardens. It is proposed to give a brief sketch of the more prominent features of the flora of the three western regions, derived partly from personal observation during a summer spent in the country, largely supplemented by the writings of the late Professor Asa Gray and other American and English botanists.
The Plains and Prairies.
The first region to be considered, that of the prairies, the great plains, and the deserts of the inland basin, may be very briefly noticed, since, although of considerable interest to the botanist, it is only occasionally that plants, remarkable for beauty of flower or other conspicuous characteristics, are met with. The eastern portion of the district, where the rich prairie lands of Kansas and Nebraska are being rapidly cultivated, produces many fine flowering plants wherever some steep or rocky slope has escaped cultivation. Here we find abundance of yuccas intermingled with blue, pink and white-flowered spider-worts, handsome large-flowered penstemons, baptisias with large pea-like flowers of blue, yellow or white, many species of astragalus, yellow and white evening-primroses and other allied forms, several cactuses of the genera opuntia and mammillaria, blue larkspurs, pink oxalises, the purple Phlox divaricata, mallows of the genera Malvastrum and Callirhoe, some of which are well-known garden plants, and a host of sunflowers, asters, cone-flowers, golden-rods, coreopsis, and many other showy composites. This is the region of the buffalo or bunch grasses which formed the chief subsistence of the American bison. They are fine-tufted bluish grasses, much resembling in appearance our fine-leaved bent grass (Agrostis setacea), which is common on the heaths about Bournemouth and in Dorsetshire. I was informed that since the bisons had been destroyed the buffalo grass was also disappearing, being replaced by various coarser growing plants and grasses. It is probable that the uniform hardening of the surface by the tread of the herds of bison, together with the equally regular manuring, favoured the growth of this particular form of grasses.
As we travel westwards, towards the Rocky Mountains, the plains become more arid, and in places the vegetation resembles that of the deserts of the great basin. Here there are fewer conspicuous flowers, and a preponderance of dwarf creeping plants, with a few thorny bushes and some species of wormwood, forming the well-known "sage-
brush" of the deserts. In the interior plains these thorny and grey-leaved shrubs prevail, with wide tracts of bare earth often covered with saline incrustations. Here and there are found some pretty flowers, such as phloxes, alliums, phacelias, gilias, cleomes, œnotheras, and other characteristic plants; but the general aspect is that of bare soil scantily covered with a dwarf vegetation, or of low, shrubby thickets of a grey or leafless aspect, consisting mostly of plants allied to the salt-wort, orache, and sea-blite of our salt-marshes, or the goose-foot and wormwood of our waste places.
The Rocky Mountains.
We will now leave these comparatively uninteresting plains and deserts and enter on the Rocky Mountains proper, their deep cañons, their wooded slopes and valleys, and their upland pastures, rocky streams and alpine heights. The forest trees consist mainly of a few species of pines, firs, and junipers, none of them very remarkable for size or beauty, with several poplars, and a few oaks, beeches, and maples; but these rarely form continuous forests, except where the soil and other conditions are especially favourable. Almost everywhere the conifers are most prominent, and give their peculiar character of dark ever-green spiriness to the forest vegetation. The present scantiness of timber trees is no doubt partly due to the agency of man, first by starting forest fires, which rapidly clear extensive areas, and more recently by the felling of timber for building and mining, a cause which has denuded most of the valleys of their original forest trees. There are a considerable number of shrubs of the usual American types, such as sumachs, snowberries, hazels, spiræas, brambles, and roses, mostly of species common to other parts of America and of no special interest from our present point of view.
It is when we enter among the mountains and explore the valleys, cañons, and lower slopes, that we meet with a variety of new and interesting plants. Among these are some which are specially characteristic of this part of North America. The phloxes, polemoniums, and gilias, some
species of which are common in our gardens, are abundant, as are the penstemons and mimuluses, with the brilliant castilleias belonging to the same family (Scrophulariaceæ), whose crimson or scarlet bracts form one of the greatest ornaments of the higher woods and pastures. The elegant genus Phacelia is not uncommon, though its chief development is in California, and the moist valley bottoms are often blue with the well-known flowers of the bulbous camassia. A curious genus of the Polygonum family (Eriogonum) is abundant, and the yellowish-white or rosy flowers of some of the species are very pleasing. Handsome composites abound, especially the genus Erigeron, with a number of peculiar forms, while the beautiful butterfly-tulips of California (Calochortus) here make their first appearance. Lupines also are plentiful, though less so than further west, and the beautiful American cowslips (Dodecatheon), sometimes called "shooting-stars," are not unfrequent in boggy meadows.
But in addition to these more or less characteristic American types, the botanist is at once struck by the appearance of a number of European or even of British plants, and these not introduced weeds but forming an essential part of the flora. This is proved by the fact that the further we penetrate among the mountains and the higher we ascend, the more numerous become these familiar species or genera. Among the more abundant of these plants are the common yarrow (Achillea millefolium), our blue hare-bell (Campanula rotundifolia), the bistort (Polyonum bistorta), the common silver-weed of our roadsides (Potentilla anserina), and the rarer shrubby cinquefoil (P. fruticosa). In the sub-alpine and alpine districts these plants of the old world become more frequent and occupy a larger space in the entire vegetation, and in order to show the importance of this interesting feature of the Rocky Mountain flora it may be well to give a brief account of a week's exploration of the vicinity of Gray's Peak, one of the highest mountains of Colorado.
Alpine Plants of the Rockies.
Accompanied by a botanical friend from Denver I went first by rail up Clear Creek Cañon, passing by Georgetown, to Graymount, the terminus of the railway, where there is an hotel and where horses are obtained for the ascent of Gray's Peak, about nine miles distant by the road. Graymount is situated at the junction of two valleys and is about 9,500 feet above the sea level. During a short stroll on the afternoon of our arrival on some rocky slopes we found two of our rarer British plants, the winter green (Pyrola rotundifolia) and the elegant twin-flower (Linnæa borealis), but instead of having nearly white flowers the former was reddish and the latter was of a deeper colour than in our native plant. The next day we walked to Kelso's cabin, where are some miners' houses about 11,000 feet above sea-level, situated at the lower end of a fine upland valley, which is above the timber line. During the earlier part of our walk up a wooded valley we first noticed fine clumps of the Siberian lungwort (Mertensia siberica) with its lovely pale blue flowers, growing more compactly than in our gardens, and splendid masses of the shrubby cinquefoil covered with its handsome yellow flowers, as well as our common harebell, all in the greatest luxuriance and beauty. In damp shady places we found the little moschatel, and in bogs the curious Swertia perennis, a kind of gentian with slaty-blue flowers. These are all European or North Asian plants, but there were many others peculiar to the region, though sometimes of European rather than American affinity. Such are the lovely columbine (Aquilegia cœrulea), allied to the species of the European Alps, abundant and conspicuous with its large blue and white flowers, while mingled with it grew the gaudy Castilleia integra, whose leafy bracts of intense crimson are visible from a long distance. This is a true American type, as is the pretty liliaceous plant, Zygadenus glaucus; and there were also abundance of dark purple or bright blue penstemons, several showy groundsels and erigerons and the handsome yellow composite, Arnica cordifolia.
It was when we had passed the timber line at about 11,500 feet elevation, and had entered the bare rocky valley at the head of which rises the snow-flecked summit of Gray's Peak, that we discovered some of the chief gems of the alpine flora of the Rocky Mountains. Along the borders of the stream, fed by the still melting snows and with its roots in the water, were fine clumps of the handsomest American primrose (Primula Parryi), its whorled flowers of a crimson-purple colour with a yellow eye resembling in general appearance the well-known Japanese primrose of our gardens. Among the stony débris and loose boulders which bordered the stream the beautiful Phacelia sericea was abundant, its violet-blue flowers growing in dense clusters and producing a charming effect among its desert surroundings. This is a typical American plant, since not only is the genus a peculiar one but the natural order to which it belongs—the Hydrophyllaceæ—is almost confined to that continent. The beautiful nemo-philas of our gardens belong to the same family. In boggy places the handsome Greenland lousewort, an Arctic species, was plentiful, and in rocky crevices we found the moss campion (Silene acaulis), which is abundant on the Scotch and Welsh mountains.
The next morning we fortunately determined to explore a lateral valley called Grizzly Gulch, which diverged to the north a mile above the hotel and led into a fine upland valley on the north side of Gray's Peak. Here, just below the timber-line, we found a miner's house, and the two miners who had come home to dinner invited us to join them, and then offered to show us a fine place for flowers. They took us through the wood for half a mile, when we came upon a rocky and grassy slope with great snow-patches in the shady hollows, and the ground which the snow had left was literally starred with flowers. Leaving us to go to their work in a mine on the steep side of the mountain, we luxuriated in the finest Alpine flower-garden we had yet seen, although my friend had visited the mountains several times. What first attracted our notice were three plants of the crowsfoot family, which grew intermingled on a grassy slope almost surrounded by
snow. These were, a nearly white globe-flower (Trollius albiflorus), very dwarf and with spreading, not globular flowers; a buttercup, whose flowers were of the most perfect circular outline, and of a pure and rich yellow, both peculiar to the Rocky Mountains; and the narcissus-flowered anemone of the European Alps. Going a little further we found some of the more characteristic American forms, such as the beautiful blue-flowered Mertensia alpina, a dwarf Alpine form of Mertensia siberica and perhaps the most lovely plant of the genus; the pretty fringed grass of parnassus (Parnassia fimbriata); with peculiar species of the European genera, Aster, Cardamine, Astragalus, Delphinium, Trifolium, Saxifraga, Sedum, Valeriana, Veronica, and Pedicularis; with others of the American genera, Phacelia, Chionophila, Mimulus, and Zygadenus; and hidden among the rocks the minute purple-flowered Primula augustifolia. What more especially interested me, however, was the number of identical British or European species. Such were the moss-campion, the Dryas octopetala, Sibbaldia procumbens, the rosewort (Sedum rhodiola), the Alpine Veronica, and the Alpine chickweed, Lloydia serotina, a small liliaceous plant found on Snowdon, and two saxifrages, Saxifraga nivalis and S. cernua, all found also in our Welsh or Scotch mountains; and of European Alpines the pretty slaty-blue Swertia perennis, which dotted the grassy slopes with its delicate flowers, the Alpine Astragalus, the Arctic willow, several saxifrages and gentians, and some other species characteristic of the flora of the Alps.
The next day, after sleeping at a miner's cabin situated at the head of the main valley at about 12,500 feet elevation, we ascended to the top of Gray's Peak, which is 14,250 feet high, and met with many other interesting plants. The little Eritrichium nanum, a minute but intensely blue forget-me-not, was found growing in the midst of clumps of the moss-campion; the Gentiana tenella and Campanula uniflora of the Arctic regions were also found at about 13,000 feet elevation; with the British Alpine penny-cress, the yellow Iceland poppy, the two-flowered sandwort, the Alpine arnica, the snowy buttercup,
and other truly Arctic plants. Along with these were a few American alpine types, such as Eriogonums, Castilleias, and several composites. Near the summit of the mountain there were alternate upward-sloping bands of loose rock-débris and short turf, the latter gay with pretty yellow flowers. On examination these were found to consist of a potentilla and a saxifrage, whose flowers, resting close on the ground, were so much alike in size, form, and colour that at a short distance they appeared identical. The intermixture of two very distinct species of flowers, coloured and shaped alike and flowering at the same time, is very uncommon, because it would interfere with regular cross-fertilization by insects. In this high and exposed situation, however, where the flowering season is very short and insects very scarce, the combination of two species of flowers may lead to a more conspicuous display, and be more attractive to whatever insects may visit such great altitudes; while with plants of such distinct families, the intermixture of the pollen would lead to no evil result, since each would be totally inert on the stigma of a flower of the other kind. The two species appear to be Saxifraga chrysantha and Potentilla dissecta.
On a general summary of the plants noticed during this excursion to one of the richest districts in the Rocky Mountains, I find that they comprised no less than 20 British species, about 45 European, mostly high Alpine or Arctic, and about 30 species which, though distinct, were allied to European types. There were thus a total of 95 species, either identical with or allied to European plants, while those which belonged to American genera, or were most nearly allied to American species, were about 30 in number. It thus appears that the alpine flora of the Rocky Mountains is mainly identical with that of the Arctic regions, and it is this identity which leads to the occurrence of so many British species in this remote district. In the review of the entire alpine flora of the Rocky Mountains by Professor Asa Gray and Sir Joseph Hooker, the number of species identical with those of the Arctic regions is 102, and the distinct, though often allied, species 81, while those that be-
long to quite peculiar American genera are only 14 in number.
In considering how this curious similarity of the alpine species of the two continents has been brought about, we must go back to a time anterior to the glacial epoch, when a rather mild climate prevailed in much of what is now the Arctic regions. The present Arctic flora, or its immediate ancestors, was then probably confined to the highest latitudes around the North Pole, together with the higher mountains which were immediately contiguous—such as Greenland, then only partially or not at all ice-clad, Spitzbergen and Nova Zembla, and some of the mountain peaks of Alaska and North-Eastern Asia. At this time the Rocky Mountains, the European Alps, and even Scandinavia supported in all probability only alpine forms of the plants of the surrounding lowlands, such as are now everywhere intermingled with the widespread Arctic species. As the cold came on, and the ice-sheet crept farther and farther over the two continents, the true Arctic plants were driven southward, displacing the indigenous flora, which could not withstand the increasing severity of the climate, and occupying all the great mountain ranges on the lower side of the ice-fields and glaciers, and also such of the peaks as rose permanently above the ice-sheet of the glacial epoch. As the cold period gradually passed away, these hardy plants kept close to the gradually retreating ice, and in this way mounted to the higher peaks of many mountains from which the ice and even perpetual snow wholly passed away. Thus it is that so many species are now common to the Rocky Mountains and the European Alps; and, what seems more extraordinary, that identical plants occur on the summits of the isolated Scotch and Welsh mountains, and also on the White Mountains of New Hampshire and some of the mountains to the south of them.
Before passing on to sketch the flora of the west coast of America, we may briefly notice the more prominent differences between the Rocky Mountain flora and that of our European Alps, such differences as must strike
every traveller who takes an interest in the floral beauties of the two regions. In the Alps the more striking and showy flowers of the Alpine pastures and higher rocks are the white, purple, and yellow anemones; the beautiful violas; the glorious blue gentians starring the short turf with azure and indigo, the numerous saxifrages, often with large and showy sprays of flowers; the many beautiful rosy and purple primulas and yellow auriculas; the handsome pinks; the delicate campanulas; the showy white and yellow buttercups, and the graceful meadow-rues. Now in almost all these groups the Rocky Mountain alpine and sub-alpine flora is deficient. Anemones are comparatively few in species and not abundant; violas are almost absent in the higher regions; gentians, though fairly abundant in species, make no brilliant display as they do in the Alps; saxifrages are few, and those of the crusted section with rigid leaves and large racemes of flowers are entirely wanting; primulas are represented by one handsome and two small and rather scarce species; campanulas are scarce, and pinks are entirely absent; while buttercups and meadow-rues are by no means abundant. Instead of these flowers so familiar to the Alpine tourists, the most showy and widespread plants are the fine and long-spurred blue-and-white columbine, and the scarlet or crimson-bracted castilleias, which form sheets of beautifully contrasted colours, often covering wide mountain slopes either above or just below the timber-line; numerous purple or blue penstemons; fine blue polemoniums and lungworts of the genus Mertensia; some handsome purple or whitish louseworts, and a host of showy purple or yellow composites, which are far more numerous and varied than in the European Alps, and occupy a more prominent place in the alpine and especially in the sub-alpine Rocky Mountain flora. It is evident, therefore, that, notwithstanding the identity of so many of the species and genera of the two regions the proportions in which they occur are very different, and the aspect of the two floras is thus altogether distinct, and in some respects strikingly contrasted.
Flora of the Sierra Nevada and California.
When we go westward to the Sierra Nevada of California, we meet with another alpine flora, generally similar to that of the Rocky Mountains, but with a smaller proportion of Arctic species and more which are characteristic of America. Here we find dwarf shrubby penstemons, curious prickly gilias, Mimulus and Eriogonum in more abundance, and a greater variety of ferns. But it is when we descend to the lower slopes and to the valleys and coast ranges of California itself that we find the greatest abundance of new plants altogether distinct from anything in the Eastern States, and it is to these that we must devote the remainder of our space.
Few countries have contributed to our gardens a larger number of showy and interesting plants than California. The rich orange yellow Eschscholtzias, the brilliant Calandrinias, the showy Godetias and Clarkias, the beautiful little Nemophilas and Phacelias, the gaudy Mimuluses and the handsome Collinsias, are known to every lover of garden flowers. Others familiar to every horticulturist are the curious pitcher-plant—Darlingtonia Californica, the handsome gigantic white poppy—Romneya coulteri, the elegant Dicentra formosa, the fine yellow-flowered shrub Fremontia Californica, the ornamental blue or white-flowered evergreens of the genus Ceanothus, the fine shrubby lupines, the lovely flowering currants, including the fine Ribes speciosum with drooping fuchsia-like flowers, the scarlet-flowered Zauschneria Californica, the fine shrubby Diplacus glutinosus, and lastly, the many ornamental bulbous plants, such as the triteleias, brodiæas, lilies, and especially the lovely butterfly-tulips of the genus Calochortus, whose flowers are most exquisitely marked inside with delicately-coloured hairy fringes. But this by no means gives an idea of the great peculiarity of the Californian flora, which is best shown by the large number of its genera, probably more than a hundred, which are altogether unknown in the Eastern States. The flora is in fact related to that of Mexico, just as the flora of the Rocky Mountains is related
to that of the Arctic regions, and the Eastern States flora to that of Japan and Eastern Asia.
But although the valleys and lowlands of California are specially characterised by hosts of brilliant annuals, monkey-flowers, lupines, and flowering shrubs, which make the country a veritable flower-garden in early spring, it is from its mountain forests of coniferæ that it derives its grandest and best-known characteristics. To a brief sketch of these, and of the accompanying shrubby and herbaceous vegetation, the remainder of this chapter will be devoted.
The Californian Forests.
The Sierra Nevada of California, though rising to nearly the same altitudes as the Rocky Mountains, is by no means an imposing range, owing to the exceedingly gradual slope of the foothills which are continuous with it. From these low and arid hills, rising with a very moderate slope from the great central valley of California, there is a constant rise over an undulating or rugged country for nearly a hundred miles to the summits of the great range. The intervening tract is often cut into deep winding valleys, whose higher slopes are terminated by rugged volcanic precipices, where they have cut through the old lava-streams that once covered a large portion of the mountains; while nearer to the crest are enormously deep valleys, bounded with vertical walls and gigantic domes or splintered peaks of granitic rocks, of which the celebrated Yosemite Valley is the best known example. Owing to this formation the summits of the range can only be seen from great distances and from a few favourable points, as a somewhat jagged line on the far horizon, just rising above the dark forest-clad slopes, and here and there flecked with perpetual snows. A coach drive of three days from the railway terminus at Milton to the Yosemite Valley, and another to the Calaveras groves of "big trees," gave me an excellent opportunity of observing the main features of this remarkable forest region.
The lower portion of the foothills up to two or three thousand feet has been greatly defaced by gold-miners,
who have dug over miles of ground and cleared away most of the fine timber. This lower portion is, however, naturally more arid, and the trees have never been so fine as at greater elevations. It is curious to notice how the pines and firs increase in beauty as well as in size as we ascend further towards the central ranges. For the first thousand feet there is a scanty vegetation of stunted shrubs, and the only conifer is the scrub-pine (Pinus sabiniana), which has a most singular appearance, being irregularly branched, with scanty foliage, and, when well grown, looking at a distance more like a poplar than a pine. Higher up occurs the large white pine (Pinus ponderosa), which, except in very fine specimens, is a coarse, unornamental tree. Above two thousand feet we meet with the sugar pine (Pinus lambertiana), so called because its turpentine is sweet and sometimes almost like a mixture of sugar and turpentine. This is a handsomer species, and when full grown is of immense size, and may be known at a distance by its clusters of large cones hanging down from the very extremities of its loftiest branches. Thus far the forests are poor, owing to the absence of the more elegant firs and cedars, which only appear above 2,500 feet, when we first meet with the noble Douglas fir and the beautiful Red cedar (Libocedrus decurrens). This last is usually known in our gardens as Thuja gigantea, characterised by its columnar mode of growth, and here sometimes reaching a hundred and fifty feet in height. Higher still, at about 4,000 feet, we come upon the most beautiful of the Californian firs, Abies concolor and A. nobilis. Both are exquisitely symmetrical in growth, while the dense horizontal branches of the latter species are adorned with the most delicate blue-green tints. These beautiful trees are to be seen here in every stage of growth, from such small plants as we see on the lawn of a suburban villa up to noble specimens 150 or 200 feet in height. These two elegant firs, along with the stately cedar and Douglas fir, and the noble yellow pine and sugar-pine, constitute the main bulk of the forest from 4,000 to 7,000 feet elevation, the belt in which alone are found the true "big trees" (Sequoia gigantea), in this country commonly known as the Wellingtonia.
VOL. I. Q
Throughout these magnificent forests there is hardly any admixture of exogenous trees, and those that do occur only form an undergrowth to the far loftier coniferæ. A few small oaks and maples are sometimes seen, but more generally there is only an undergrowth of beautiful shrubs, the most conspicuous being the fine Californian dog-wood (Cornus nuttalii), whose flowers, formed of the white involucres, are six inches across; and the lovely white azalea, whose delicate blossoms are beautifully marked with yellow. Besides these are the handsome Californian laurel and the white or blue flowered Ceanothus, while the "madrono" and "manzanita" (species of Arbutus and Arctostaphylos), are found in the drier portions of the forest and at a lower elevation.
The ground under the pines and firs is usually rather bare, but in favourable places there are some curious or beautiful creeping or herbaceous plants. Some of the drier slopes are completely carpeted with a curious little rosaceous plant (Chamæbatia foliolosa), having white flowers like those of a bramble and the most minutely divided tripinnate foliage, each leaflet looking about the size of a pin's head. Perhaps the most remarkable herbaceous plant of these forests is the Sarcodes sanguinea, a leafless parasite allied to our native monotropa, but of an intense crimson colour and very large, being often more than a foot high and two or three inches diameter. It is called the "snow-plant" in California, because it appears before the snow has wholly melted and is most striking and beautiful when growing out of it. This plant is accurately represented in one of the pictures in the "North" gallery at Kew. On the sides of the rocky streams growing in fissures which are often under water, the large peltate saxifrage seems quite at home, although in our gardens it will grow and flower even in the driest situations. The fine shrubby Penstemon newberryi also adorns the rocky margins of the streams, the beautiful Diplacus glutinosus of our greenhouses is a common wayside shrub, while the lovely blue Brodiæas and painted Calochorti or butterfly-tulips, are as common as our bluebells and poppies. The fine yellow Cypripedium montanum is occasionally found
in the forest bogs, while in open ground near the "Big Tree" Hotel, exquisite little blue Nemophilas, yellow Mimulus, and a tall Echinospermum with flowers like a large forget-me-not, were very abundant. Among these and many other strange flowers one British species was found, often starring the ground under the giant trees with its delicate flowers. This was the little chickweed winter-green (Trientalis Europæa), only differing from our native plant in the flowers being pale pink instead of white.
Even if we leave out of consideration the giant Sequoias, the forests of the Sierra Nevada would stand pre-eminent for the beauty and grandeur of their pines, firs, and cedars. Three of these, the white pine, the red cedar, and the sugar-pine are, not unfrequently, more than six feet in diameter at five or six feet above the ground, whence the giant trunks taper very gradually upwards. One sugar-pine near the Big-Tree Hotel was found to be seven feet two inches in diameter at five feet above the ground. A red cedar measured at the same height was seven feet diameter, and one of the white pines five feet nine inches. The height of the above-named sugar-pine was measured approximately by means of its shadow, and found to be 225 feet, and I was assured that one which had been cut down near the hotel was 252 feet high. The Douglas fir in the forest of British Columbia is said to surpass these dimensions considerably, being often ten feet or even twelve feet diameter, and near 300 feet high. Probably in no other part of the world than the west coast of North America is there such a magnificent group of trees as these; yet they are all far exceeded by two others inhabiting the same country, the two Sequoias—S. gigantea and S. sempervirens.
The Big Trees of California.
In the popular accounts of these trees it is usual to dwell only on the dimensions of the very largest known specimens, and sometimes even to exaggerate these. Even the smaller full-grown trees, however, are of grand
Q 2
dimensions, varying from 14 to 18 feet in diameter at six feet above the ground, and keeping nearly the same thickness for perhaps a hundred feet. In the south Calaveras grove, where there are more than a thousand trees, the exquisite beauty of the trunks is well displayed by the numerous specimens in perfect health and vigour. The bark of these trees, seen at a little distance, is of a bright orange brown tint, delicately mottled with darker shades, and with a curious silky or plush-like gloss, which gives them a richness of colour far beyond that of any other conifer. The tree which was cut down soon after the first discovery of the species, the stump of which is now covered with a pavilion, is 25 feet in diameter at six feet above the ground, but this is without the thick bark, which would bring it to 27 feet when alive. A considerable portion of this tree still lies where it fell, and at 130 feet from the base I found it to be still 12½ feet in diameter (or 14 feet with the bark), while at the extremity of the last piece remaining, 215 feet from its base, it is six feet in diameter, or at least seven feet with the bark. The height of this tree when it was cut down is not recorded, but as one of the living trees is more than 360 feet high, it is probable that this giant was not much short of 400 feet.
In the accompanying photo-plate the dead tree in the centre is that from which the bark was stripped which was erected in the Crystal Palace and unfortunately destroyed by fire. It is called the "Mother of the Forest." The two trees nearer the foreground are healthy, medium-sized trees, about 15 feet diameter at 6 feet above the ground.
The huge decayed trunk called "Father of the Forest," which has fallen perhaps a century or more, exhibits the grandest dimensions of any known tree. By measuring its remains, and allowing for the probable thickness of the bark, it seems to have been about 35 feet diameter near the ground, at 90 feet up to 15 feet, and even at a height of 270 feet it was 9 feet diameter. It is within the hollow trunk of this tree that a man on horseback can ride— both man and horse being rather small; but the dimensions undoubtedly show that it was considerably larger than the "Pavilion tree," and that it carried its huge

FIG. 49.—THE "MOTHER OF THE FOREST" (DEAD) CALAVERAS GROVE.
dimensions to a greater altitude; and although this does not prove it to have been much taller, yet it was in all probability more than 400 feet in height.
Very absurd statements are made to visitors as to the antiquity of these trees, three or four thousand years being usually given as their age. This is founded on the fact that while many of the large Sequoias are greatly damaged by fire the large pines and firs around them are quite uninjured. As many of these pines are assumed to be near a thousand years old, the epoch of the "great fire" is supposed to be earlier still, and as the Sequoias have have not outgrown the fire-scars in all that time they are supposed to have then arrived at their full growth. But the simple explanation of these trees alone having suffered so much from fire is, that their bark is unusually thick, dry, soft, and fibrous, and it thus catches fire more easily and burns more readily and for a longer time than that of the other coniferæ. Forest fires occur continually, and the visible damage done to these trees has probably all occurred in the present century. Professor C. B. Bradley, of the University of California, has carefully counted the rings of annual growth on the stump of the "Pavilion tree," and found them to be 1,240; and after considering all that has been alleged as to the uncertainty of this mode of estimating the age of a tree, he believes that in the climate of California, in the zone of altitude where these trees grow, the seasons of growth and repose are so strongly marked that the number of annual rings gives an accurate result.
Other points that have been studied by Professor Bradley are, the reason why there are so few young trees in the groves, and what is the cause of the destruction of the old trees. To take the last point first, these noble trees seem to be singularly free from disease or from decay due to old age. All the trees that have been cut down are solid to the heart, and none of the standing trees show any indications of natural decay. The only apparent cause for their overthrow is the wind, and by noting the direction of a large number of fallen trees it is found that the great majority of them lie more or less towards the south. This is not the direction of the prevalent winds, but many
of the tallest trees lean towards the south, owing to the increased growth of their topmost branches towards the sun. They are then acted upon by violent gales, which loosen their roots, and whatever the direction of the wind that finally overthrows them, they fall in the direction of the overbalancing top weight. The young trees grow spiry and perfectly upright, but so soon as they overtop the surrounding trees and get the full influence of the sun and wind, the highest branches grow out laterally, killing those beneath by their shade, and thus a dome-shaped top is produced. Taking into consideration the health and vigour of the largest trees, it seems probable that, under favourable conditions of shelter from violent winds and from a number of trees around them of nearly equal height, big trees might be produced far surpassing in height and bulk any that have yet been discovered. It is to be hoped that if any such are found to exist in the extensive groves of these trees to the south of those which are alone accessible to tourists, the Californian Government will take steps to reserve a considerable tract containing them, for the instruction and delight of future generations.
The scarcity of young Sequoias strikes every visitor, the fact being that they are only to be found in certain favoured spots. These are, either where the loose debris of leaves and branches which covers the ground has been cleared away by fire, or on the spots where trees have been uprooted. Here the young trees grow in abundance, and serve to replace those that fall. The explanation of this is, that during the long summer drought the loose surface debris is so dried up that the roots of the seedling Sequoias perish before they can penetrate the earth beneath. They require to germinate on the soil itself and this they are enabled to do when the earth is turned up by the fall of a tree, or where a fire has cleared off the debris. They also flourish under the shade of the huge fallen trunks in hollow places, where moisture is preserved throughout the summer. Most of the other conifers of these forests, especially the pines, have much larger seeds than the Sequoias, and the store of nourish-
ment in these more bulky seeds enables the young plants to tide over the first summer's drought. It is clear, therefore, that there are no indications of natural decay in these forest giants. In every stage of their growth they are vigorous and healthy, and they have nothing to fear except from the destroying hand of man.
Destruction from this cause is, however, rapidly diminishing both the giant Sequoia and its near ally the noble redwood (Sequoia sempervirens), a tree which is more beautiful in foliage and in some other respects more remarkable than its brother species, while there is reason to believe that under favourable conditions it reaches an equally phenomenal size. It once covered almost all the coast ranges of central and northern California, but has been long since cleared away in the vicinity of San Francisco, and greatly diminished elsewhere. A grove is preserved for the benefit of tourists near Santa Cruz, the largest tree being 296 feet high, 29 feet diameter at the ground, and 15 feet at 6 feet above it. One of these trees having a triple trunk is here figured from a photograph. Much larger trees, however, exist in the great forests of this tree in the northern part of the State, but these are rapidly being destroyed for the timber, which is so good and durable as to be in great demand. Hence Californians have a saying that the redwood is too good a tree to live. On the mountains a few miles east of the Bay of San Francisco, there are numbers of patches of young redwoods indicating where large trees have been felled, it being a peculiarity of this tree that it sends up vigorous young plants from the roots of old ones immediately around the base. Hence in the forests these trees often stand in groups arranged nearly in a circle, thus marking out the size of the huge trunks of their parents. It is from this quality that the tree has been named "sempervirens," or ever flourishing. Dr. Gibbons, of Alameda, who has explored all the remains of the redwood forests in the neighbourhood of Oakland, kindly took me to see the old burnt-out stump of the largest tree he had discovered. It is situated about 1,500 feet above the sea and is 34 feet in diameter at the ground. This

FIG. 50.—REDWOOD TREE (sequoia sempervirens) SANTA CRUZ
is as large as the very largest specimens of the Sequoia gigantea, but it may have spread out more at the base and have been somewhat smaller above, though this is not a special characteristic of the species. Many other stumps were seen which were 20 and 30 feet in diameter, and all were surrounded with young trees of various sizes. The large tree is said to have been cut down forty years ago. It is, therefore, probable that, in the forests to the northward, redwood trees may exist equalling, if not surpassing, the "big trees" themselves.
I have now concluded a very brief and imperfect sketch of the more prominent aspects of North American vegetation, as seen during a single summer's travel across the continent. Many grand and beautiful scenes remain vividly painted on my memory; but if I were asked what most powerfully impressed me, as at once the grandest and most interesting of the many wonders of the Western world, I should answer, without hesitation, that it was the two majestic trees, some account of which I have just given, together with the magnificent and beautiful forests in the heart of which they are found. Neither the thundering waters of Niagara, nor the sublime precipices and cascades of Yosemite, nor the vast expanse of the prairies, nor the exquisite delight of the alpine flora of the Rocky Mountains—none of these seem to me so unique in their grandeur, so impressive in their display of the organic forces of nature, as the two magnificent "big trees" of California. Unfortunately these alone are within the power of man totally to destroy, as they have been already partially destroyed. Let us hope that the progress of true education will so develop the love and admiration of nature, that the possession of these altogether unequalled trees will be looked upon as a trust for all future generations, and that care will be taken, before it is too late, to preserve not only one or two small patches, but some more extensive tracts of forest, in which they may continue to flourish, in their fullest perfection and beauty, for thousands of years to come, as they have flourished in the past, in all probability for millions of years and over a far wider area.
CHAPTER XI
THE PALæARCTIC AND NEARCTIC REGIONS COMPARED AS REGARDS THE FAMILIES AND GENERA OF THEIR MAMMALIA AND BIRDS
IN a paper read before the Cambridge Natural Science Club on March 12 (and printed in Nature of April 26, 1894), I discussed the question of the nature and uses of Zoological Regions, and arrived at the conclusion that, in order to secure the maximum of utility, it is essential to have only one set of regions for all groups of land animals; and, further, that the six regions established by Dr. P. L. Sclater are the most natural and convenient, and are best adapted to facilitate the comparative study of distribution, which is the main purpose for which Zoological Regions, as distinct from the ordinary geographical divisions of the globe, have been established.
These regions were at first generally adopted; but of late years many eminent naturalists, both in America and Europe, have proposed other divisions, though hardly any two of these agree with each other. The most important modification, and that which has been adopted by several zoologists both as regards mammals and birds, is to unite the Palæarctic and Nearctic Regions so as to form one new region, coextensive with the extratropical Northern Hemisphere. This new region, which has been termed the Holarctic, is said to be more nearly equal to the other regions as regards peculiar genera and families, and, therefore, to form part of a more natural and harmonious series than if we treat the two component parts as
separate regions. It is this one question only that I propose to discuss in the present chapter; whether the Palæarctic and Nearctic Regions, as defined by Sclater, present so many resemblances and so few differences that they can be reasonably considered to form one region as homogeneous as are most of the other regions.
The reason why so much difference of opinion exists on this point, when the very same facts are before all the enquirers, seems to be that they treat the facts in different ways. In the first place, it appears to me that far too much stress is laid upon the comparatively small number of absolutely peculiar genera or families in the two temperate regions. That may, or may not, be a reasonable ground for deciding that the regions in question are not of equal rank with some other regions; but it cannot justify the union of the two areas unless it can be shown that they are to a large extent homogeneous— that, in fact, the resemblances are more numerous and more important than the differences. In the areas we are discussing, however, this is decidedly not the case, since the facts clearly show that the differences very far surpass the resemblances—that the differences are, in fact, fundamental, and are far greater than can be found in the separate halves of any of the other regions, unless they are so divided as to be very unequal in area or to present very great diversity of climate. But the Palæarctic and Nearctic Regions are, roughly speaking, equal in area, while they both exhibit the very same range of climate. Any difference that exists between them must, therefore, be due to more fundamental causes; and the most fundamental cause is, that in each case the existing fauna is descended from an ancient, equally distinct, fauna, modified in different ways by immigration from adjacent areas.
In dealing with any question of this kind, it is very important that the genera and family groups adopted shall be the same as far as possible throughout the whole area. In order to ensure this, I have, for mammalia, adopted those of Flower and Lydekker in their Mammals Living
and Extinct. In the case of the birds, I am indebted to my friend Mr. H. E. Dresser, both for completing his List of European Birds so as to include those of the whole Palæarctic region, and also for so harmonising his genera with those of the A. O. U. Check List of North American Birds as to combine the whole into a single series. As Mr. Dresser has made a study of North American birds as well as of those of the Palæarctic Region with which his name is more especially associated, the result is probably as fair and unbiassed, for the comparison in question, as can be arrived at. In the case of both regions, I have rejected all those genera in which a single species only just enters the region from the adjacent region to which it properly belongs. Here, again, there is, of course, room for difference of opinion; but the doubtful cases are not sufficiently numerous to introduce any important uncertainty into the result.
For both mammals and birds I adopt the same system of presenting the facts which, after much consideration and many trials, seems to me to be the best. The families and genera are arranged in three columns—the first giving those which are Palæarctic but not Nearctic; the third those that are Nearctic but not Palæarctic; the middle column giving those which are common to the two regions —so that by combining this with either of the others we obtain the entire fauna of that region. The results are then summarised in other tables and briefly discussed. We begin with the mammals:—
MAMMALS OF THE PALæARCTIC AND NEARCTIC REGIONS.
| PALæARCTIC. | BOTH REGIONS. | NEARCTIC. |
| BATS. | ||
| RHINOLOPHIDæ (Horseshoe Bats): | ||
| 1. Rhinolophus. | ||
| VESPERTILIONIDæ (Bats): | VESPERTILIONIDæ: | VESPERTILIONIDæ: |
| 2. Synotus. | 1. Plecotus. | 1. Antrozous. |
| 3. Harpyiocephalus. | 2. Vesperugo. | 2. Nycticejus. |
| 3. Vespertilio. | 3. Atalapha. | |
| EMBALLONURIDæ: | PHYLLOSTOMIDæ: | |
| 4. Nyctinomus. | 4. Macrotus. |
| PALæARCTIC. | BOTH REGIONS. | NEARCTIC. |
| INSECTIVORA. | ||
| ERINACEIDæ (Hedgehogs): | ||
| 4. Erinaceus. | ||
| SORICIDæ (Shrews): | SORICIDæ: | SORICIDæ: |
| 5. Crossopus. | 5. Sorex. | 5. Neosorex. |
| 6. Crocidura. | 6. Blarina. | |
| 7. Diplomesodon. | ||
| 8. Anurosorex. | ||
| 9. Nectogale. | ||
| TALPIDæ (Moles): | TALPIDæ: | TALPIDæ: |
| 10. Myogale. | 6. Urotrichus. | 7. Scalops. |
| 11. Talpa. | 8. Scapanus. | |
| 9. Condylura. | ||
| CARNIVORA. | ||
| VIVERRIDæ (Civets): | FELIDæ (Cats). | PROCYONIDæ (Racoons): |
| 12. Genetta. | 7. Felis. | 10. Procyon. |
| 13. Herpestes. | 11. Bassaris. | |
| HYæNIDæ (Hyænas): | CANIDæ (Wolves): | |
| 14. Hyæna. | 8. Canis. | |
| URSIDæ (Bears): | ||
| 9.Ursus. | ||
| MUSTELIDæ (Weasels): | MUSTELIDæ: | MUSTELIDæ: |
| 15. Meles (Badger). | 10. Lutra (Otter). | 12. Latax. |
| 11. Mustela. | 13. Mephitis (Skunk). | |
| 12. Gulo (Glutton). | 14. Taxidea. | |
| TRICHECHIDæ (Walrus): | OTARIIDæ (Eared Seals): | |
| 13. Trichechus. | 15. Otaria. | |
| PHOCIDæ (Seals): | PHOCIDæ: | PHOCIDæ: |
| 16. Halichærus. | 14. Phoca. | 16. Macrorhinus. |
| 17. Monachus. | 15. Cystophora. | |
| UNGULATA. | ||
| SUIDæ (swine): | DICOTYLIDæ (Peccaries): | |
| 18. Sus. | 17. Dicotyles. | |
| CERVIDæ (Deer): | CERVIDæ: | CERVIDæ: |
| 19. Moschus (Musk deer). | 16. Cervus. | 18. Cariacus. |
| 17. Rangifer (Reindeer). | ||
| 18. Alces. | ||
| ANTILOCAPRIDæ: | ||
| 19. Antilocapra (pronghorn). | ||
| BOVIDæ (Antelopes): | BOVIDæ: | BOVIDæ: |
| 20. Saiga. | 19. Ovis (Sheep). | 20. Haplocerus (American Antelope). |
| 21. Pantholops. | 20. Bos (Oxen). | |
| 22. Gazella. | 21. Ovibos (Musk-Sheep). | |
| 23. Addax. | ||
| 24. Rupicapra. | ||
| 25. Budorcas. | ||
| 26 Capra. | ||
| EQUIDæ (Horses): | ||
| 27. Equus. |
| PALæARCTIC. | BOTH REGIONS. | NEARCTIC. |
| UNGULATA—(continued): | ||
| RODENTIA. | ||
| SCIURIDæ (Squirrels): | SCIURIDæ: | |
| 21. Sciurus. | 22. Cynomys. | |
| 22. Tamias. | ||
| 23. Sciuropterus. | ||
| 24. Arctomys(Marmots). | ||
| 25. Spermophilus. | ||
| MYOXIDæ (Dormice): | CASTORIDæ (Beavers): | HAPLOODONTIDæ: |
| 28. Myoxus. | 26. Castor. | 23. Haploodon. |
| 29. Eliomys. | ||
| 30.Muscardinus. | ||
| MURIDæ (Mice, &c.): | MURIDæ: | MURIDæ: |
| 31. Gerbillus. | 27. Arvicola. | 24. Hesperomys. |
| 32. Cricetus. | 28. Myodes. | 25. Sigmodon. |
| 33. Ellobius. | 29. Cuniculus. | 26. Ochetodon. |
| 34. Siphneus. | 27. Neotoma. | |
| 35. Mus. | 28. Phenacomys. | |
| 36. Acanthomys. | 29. Synaptomys. | |
| 30. Fiber. | ||
| 31. Neofiber. | ||
| SPALACIDæ (Blind Rats): | ||
| 37. Spalax. | ||
| 38. Rhizomys. | GEOMYIDæ (Pouched Rats): | |
| 32. Geomys. | ||
| 33. Thomomys. | ||
| 34. Diplodomys. | ||
| 35. Perognathus. | ||
| 36. Heteromys. | ||
| DIPODIDæ (Gerboas): | DIPODIDæ: | |
| 39. Sminthus. | 37. Zapus. | |
| 40. Dipus. | ||
| 41. Alactaga. | ||
| 42. Platyceromys. | ||
| HYSTRICIDæ (Porcupine): | HYSTRICIDæ: | |
| 43. Hystrix. | 38. Erethizon (Prehensile tailed Porcupine). | |
| LAGOMYIDæ (Pikas): | ||
| 30. Lagomys. | ||
| LEPORIDæ(Hares): | ||
| 31. Lepus. | ||
| MARSUPIALIA. | ||
| DIDELPHIDæ (Opossums): | ||
| 39. Didelphys. |
Looking through the preceding lists, we find a remarkable divergence in the characteristic groups of the two regions. Out of fifteen genera of Insectivora, only two are common to both regions; an amount of difference which, if it occurred among larger and better known animals, would produce a striking effect of diversity. In the Car-
nivora there is a much smaller proportion of peculiar genera, but this is to some extent counterbalanced by each region possessing two families absent from the other. Ungulata, again, show considerable diversity, the Palæarctic Region possessing ten genera and the Nearctic five which are not found in the other. Only five genera are common to both, and of these five, two, Rangifer and Alces, are arctic, while two others, Ovis and Bos, have one species each in the Nearctic Region against about thirteen between them in the Palæarctic. The Rodents, again, have only eleven genera common to both regions, while the Palæarctic has sixteen and the Nearctic seventeen which are not found in the other region.
The following summary will enable us to see the total amount of similarity and difference:—
SUMMARY OF DISTRIBUTION.
| ORDERS. | FAMILIES. | GENERA. | ||||||
| No. of Genera. | Palæarctic. | Both. | Nearctic. | Palæarctic. | Both. | Nearctic. | ||
| Cheiroptera | 11 | 1 | 2 | 1 | 3 | 4 | 4 | |
| Insectivora | 15 | 1 | 2 | 0 | 8 | 2 | 5 | |
| Carnivora | 22 | 2 | 6 | 2 | 6 | 9 | 7 | |
| Ungulata | 20 | 2 | 2 | 2 | 10 | 5 | 5 | |
| Rodentia | 44 | 2 | 7 | 2 | 16 | 11 | 17 | |
| Marsupialia | 1 | 0 | 0 | 1 | 0 | 0 | 1 | |
| 8 | 19 | 8 | 43 | 31 | 39 | |||
| Total Genera | 113 | — | — | — | 74 | — | 70 | |
| Total Families | 27 | 27 |
Here we see that in both the regions the number of genera not found in the other largely exceed those common to both, the proportion of the total genera thus limited being 58 per cent. in the Palæarctic, and 56 per cent. in the Nearctic Region; while in each case out of a total of 27 families no less than 8 are so limited. If we compare this amount of diversity with that between the Ethiopian and Oriental Regions, we shall find that, while as regards genera it is somewhat less, as regards families it is considerably greater. In the Oriental Region out of a total of about 72 families of mammals only 8
are not Ethiopian; while in the Ethiopian region out of about 74 families only 10 are not Oriental. No doubt these two regions are those which most resemble each other, and they have been united by some naturalists, but they are now generally admitted to be sufficiently distinct to be classed as separate regions. Great weight has, however, always been given to the possession of peculiar families; and for the same reason the fact that nearly one-third of the families in the Palæarctic and Nearctic regions respectively are not found in the other, indicates a fundamental diversity that renders their union into one region quite inadmissible.
We will now tabulate the birds, in which the amount of diversity is much larger as regards genera, and equally as large as regards families:—
LAND BIRDS OF THE PALæARCTIC AND NEARCTIC REGIONS.
| PALæARCTIC. | BOTH. | NEARCTIC. |
| TURDIDæ (Thrushes): | TURDIDæ: | TURDIDæ: |
| 1. Monticola (4 sp.). | 1. Turdus (P.21 sp. N. 5 sp.). | 1. Merula (2). |
| 2. Myiophoneus (1). | 2. Hesperocichla (1) | |
| 3. Saxicola (20). | 2. Sialia (1) (=Grandala (3). | 3. Polioptila (3). |
| 4. Pratincola (7). | 4. Myadestes (1). | |
| 5. Ruticilla (15). | 3. Regulus (P. 21 sp N. 5 sp.) (4.3). | |
| 6. Hodgsonius (1). | ||
| 7. Chimarrhornis (1). | ||
| 8. Cyanecula (2). | ||
| 9. Erithacus (4). | ||
| 10. Calliope (3). | ||
| 11. Cossypha (1). | ||
| 12. Nemura (1). | ||
| 13. Daulias (2). | ||
| 14. Sylvia (18). | ||
| 15. Melizophilus (2). | ||
| 16. Phylloscopas (19). | ||
| 17. Hypolais (8). | ||
| 18. ædon (2). | ||
| 19. Acrocephalus (10). | ||
| 20. Lusciniola (10). | ||
| 21. Locustella (8). | ||
| 22. Cettia (4). | ||
| 23. Cisticola (1) | ||
| 24. Drymoeca (1) | ||
| 25. Scotocerca (2). | ||
| 26. Argya (3). | ||
| 27. Rhopophilus (3). | ||
| 48. Babax (1). | ||
| 29. Peterorhinus (1). | ||
| 30. Trochalopteron (2). | ||
| CINCLIDæ (Dippers). | ||
| 4. Cinclus (4.1). | ||
| ACCENTORIDæ (Hedge sparrows): | ||
| 31. Accentor (11). |
VOL. I. R
| PANURIDæ (Bearded Tits): | ||
| 32. Panurus (1). | ||
| PARIDæ (Titmice): | PARIDæ: | PARIDæ: |
| 33. Leptopœcilus (2). | 5. Parus (26.6). | 5. Chamæa (12) (sub. fam). |
| 34. Lophobasileus (1). | 6. Lophophanes (3.4). | |
| 35. Suthora (1). | 7. Acredula (7) = Psaltriparus (13). | |
| 36. ægithalus (4). | ||
| SITTIDæ (Nuthatches). | ||
| 8. Sitta (8.4). | ||
| CERTHIIDæ (Creepers): | CERTHIIDæ: | |
| 37. Tichodroma (1). | 9. Certhia (2.1). | |
| TROGLODYTIDæ (Wrens): | TROGLODYTIDæ: | |
| 10. Troglodytes (6.3). | 6. Cistothorus (2). | |
| 7. Thryothorus (3). | ||
| 8. Catherpes (1). | ||
| MOTACILLIDæ (Wagtails): | MOTACILLIDæ: | 9. Salpinctes (2). |
| 38. Motacilla (16). | 11. Anthus (14.2). | 10. Harporhynchus (8). |
| 39. Limodromus (1). | 11. Galeoscoptes (1). | |
| 12. Mimus (1). | ||
| 13. Oroscoptes (1). | ||
| CRATEROPODIDæ? | MNIOTILTIDæ (Wood Warblers). | |
| 40. Zosterops (2). | ||
| 14. Mniotilta (1). | ||
| ORIOLIDæ (Orioles): | 15. Protonotaria (1). | |
| 41. Oriolus (2) | 16. Helenaia (1). | |
| 17. Helmitherus (1). | ||
| 18. Helminthophila (8) | ||
| 19. Compsothlypis (2). | ||
| 20. Dendroica (24). | ||
| 21. Sciurus (3). | ||
| 22. Geothlypis (6). | ||
| 23. Icteria (1). | ||
| 24. Sylvania (3). | ||
| 25. Setophaga (2). | ||
| 26. Cardellina (1). | ||
| MUSCICAPIDæ (Flycatchers): | LANIIDæ (Shrikes). | VIREONIDæ (Greenlets). |
| 42. Muscicapa (6). | 12. Lanius (22.2). | 27. Vireo (12). |
| 43. Hemichelidon (1). | ||
| 44. Alseonax (1). | AMPELIDæ (Waxwings): | AMPELIDæ: |
| 45. Xanthopygia (3). | 13. Ampelis (2.2). | 28. Phainopepla (1). |
| HIRUNDINIDæ (Swallows): | HIRUNDINIDæ: | HIRUNDINIDæ: |
| 46. Chelidon (3). | 14. Hirundo (3) (= Chelidon (1). | 29. Progne (1). |
| 30. Petrochelidon (1). | ||
| 15. Cotile (3) (= Clivicola (1). | 31. Tachyniceta (2). | |
| 32. Stelgidopteryx (1). | ||
| TANAGRIDæ (Tanagers): | ||
| 33. Piranga (4). | ||
| FRINGILLIDæ (Finches): | FRINGILLIDæ: | FRINGILLIDæ: |
| 47. Carduelis (2). | 16. Chrysomitrus (2)= Spinus (4) | 34. Rhyncophanes (1). |
| 48. Serinus (4). | 35. Poocetes (1). | |
| 49. Chloris (3). | 17. Coccothraustes (1.1) | 36. Ammodramus (11) |
| 50. Passer (8). | 18. Linota (7) = Acanthis (2). | 37. Chondestes (1). |
| 51. Eophona (2). | 38. Zonotrichia (6). | |
| 52. Pycnorhamphus (1). | 19. Carpodacus (10.4) | 39. Spizella (7). |
| 53. Petronia (2). | 20. Pinicola (1.1). | 40. Junco (7). |
| 54. Montifringilla (7). | 21. Loxia (5.2). | 41. Amphispiza (2). |
| FRINGILLIDæ (Finches)—(continued): | FRINGILLIDæ—(continued): | FRINGILLIDæ—(continued): |
| 55. Fringilla (6). | 22. Plectrophanes (2)= Calcarius and Plectrophenax (5). | 42. Peucæa (5). |
| 56. Fringalauda (1). | 43. Melospiza (4). | |
| 57. Bucanetes (4). | 44. Passerella (1). | |
| 58. Uragus (2). | 23. Leucosticte (5.3). | 45. Pipilo (6). |
| 59. Pyrrhula (7). | 46. Cardinalis (1). | |
| 60. Urocynchramus (1). | 47. Habia (2). | |
| 61. Emberiza (32). | 48. Guiraca (1). | |
| 49. Passerina (4). | ||
| 50. Spiza (1). | ||
| 51. Calamospiza (1). | ||
| ICTERIDæ (American Starlings): | ||
| 52. Dolichonyx (1). | ||
| 53. Molothrus (1). | ||
| 54. Xanthocephalus (1). | ||
| 55. Agelaius (3). | ||
| 56. Sturnella (1). | ||
| 57. Icterus (6). | ||
| 58. Scolecophagus (2). | ||
| 59. Quiscalus (3). | ||
| ALAUDIDæ (Larks): | ALAUDIDæ: | |
| 62. Certhilauda (2). | 24. Otocorys (4.1). | |
| 63. Galerita (3). | ||
| 64. Alauda (3). | ||
| 65. Ammomanes (2). | ||
| 66. Calandrella (6). | ||
| 67. Melanoscorypha (6). | ||
| 68. Rhamphocorys (1). | ||
| STURNIDæ (Starlings): | ||
| 69. Sturnus (4). | ||
| 70. Pastor (1). | ||
| 71. Poliopsar (1). | ||
| 72. Sturnia (2). | ||
| CORVIDæ (Crows): | CORVIDæ: | CORVIDæ: |
| 73. Podoces (4). | 25. Nucifraga (1) = Picicorvus (1). | 60. Cyanocitta (2). |
| 74. Pyrrhocorax (2). | 61. Aphelocoma (3). | |
| 75. Garrulus (9). | 26. Perisoreus (1.2). | 62. Cyanocephalus (1). |
| 76. Cyanopica (2). | 27. Pica (2.2). | |
| 28. Corvus (11.5). | ||
| TYRANNIDæ (Tyrantflycatchers): | ||
| 63.Milvulus (1). | ||
| 64. Tyrannus (4). | ||
| 65. Myiodynastes (1). | ||
| 66. Myiarchus (3). | ||
| 67. Sayornis (3). | ||
| 68. Contopus (4). | ||
| 69. Empidonax (7). | ||
| 70. Pyrocephalus (1). | ||
| 71. Ornithion (1). | ||
| CYPSELIDæ (Swifts). | CYPSELIDæ: | CYPSELIDæ: |
| 77. Acanthyllis (1). | 29. Cypselus (7) = Micropus (1). | 72. Cypseloides (1). |
| 73. Chætura (2). | ||
| CAPRIMULGIDæ (Night-jars). | CAPRIMULGIDæ: | |
| 78. Caprimulgus (4). | 74. Antrostomus (2). | |
| 75. Phalænoptilus (1). | ||
| 76. Chordeiles (1). |
R 2
| TROCHILIDæ (Hummingbirds) | ||
| 77. Eugenes (1). | ||
| 78. Cœligena (1). | ||
| 79. Trochilus (8). | ||
| 80. Iache (1). | ||
| PICIDæ (Woodpeckers): | PICIDæ: | PICIDæ: |
| 79.Dryocopus (2). | 30. Picus (18) = Dryobates (5). | 81. Campephilus (1). |
| 80. Gecinus (6). | 82.Xenopicus (1). | |
| 81. Ingipicus (3). | 31. Picoides (1.2). | 83. Sphyrapicus (3). |
| 82. Jynx (1). | 84. Ceophlæus (1). | |
| 85. Melanerpes (5). | ||
| 86. Colaptes (4). | ||
| ALCEDINIDæ (Kingfishers): | ALCEDINIDæ: | |
| 83. Alcedo (1). | 32. Ceryle (2.2). | |
| 84. Halcyon (2). | ||
| CORACIADæ (Rollers): | ||
| 85. Coracias (2). | ||
| MEROPIDæ (Bee eaters): | ||
| 86. Merops (3). | ||
| UPUPIDæ (Hoopoes): | ||
| 87. Upupa (1). | ||
| CUCULIDæ (Cuckoos): | CUCULIDæ: | |
| 88. Cuculus (4). | 87. Coccyzus (2). | |
| 89. Hierococcyx (2). | 88. Geococcyx (1). | |
| 90. Coccystes (1). | 89. Crotophaga (1). | |
| PSITTACIDæ (Parrots): | ||
| 90. Conurus (1). | ||
| STRIGIDæ (Barn owls). | ||
| 33. Strix (1.1). | ||
| BUBONIDæ (Horned owls): | BUBONIDæ: | BUBONIDæ: |
| 91. Ninox (1). | 34. Asio (1.2). | 91. Speotypo (1). |
| 92. Athene (3). | 35. Syrnium (4) = Ulula (3). | 92. Micrathene (1). |
| 36. Nyctea (1.1). | ||
| 37. Surnia (2.1). | ||
| 38. Nyctala (1.2). | ||
| 39. Scops (4) = Megascops (2). | ||
| 40. Bubo (4.1). | ||
| 41. Glaucidium (1.1). | ||
| VULTURIDæ (Vultures): | CATHARTIDæ (American Vultures): | |
| 93. Gyps (2). | ||
| 94. Vultur (1). | 93. Pseudogryphus (1). | |
| 95. Neophron (1). | 94. Cathartes (1). | |
| 96. Gypaetus (1). | 95. Catharista (1). | |
| FALCONIDæ (Falcons): | FALCONIDæ: | FALCONIDæ: |
| 97. Circaetus (1). | 42. Circus (6.1). | 96. Elanoides (1). |
| 98. Nisaetus (2). | 43. Buteo (5.7). | 97. Ictinia (1). |
| 99. Milvus (4). | 44. Archibuteo (2.2). | 98. Parabuteo (1). |
| 100. Pernis (1). | 45. Aquila (7.1). | 99. Urubutinga (1). |
| 46. Haliætus (3.1). | 100. Polyborus (2). | |
| 47. Astur (2.1). | ||
| 48. Accipiter (4.2). | ||
| 49. Elanus (1.1). | ||
| 50. Falco (17.8). | ||
| 51. Pandion (1.1). |
| COLUMBIDæ (Pigeons): | COLUMBIDæ: | COLUMBIDæ: |
| 101. Turtur (8). | 52. Columba (14.3). | 101. Ectopistes (1). |
| 102. Zenaidura (1). | ||
| 103. Columbigallina (1). | ||
| 104. Scardafella (1). | ||
| PTEROCLIDæ (Sand grouse): | ||
| 102. Pterocles (6). | ||
| 103. Syrrhaptes (2). | ||
| PHASIANIDæ (Pheasants): | PHASIANIDæ: | |
| 104. Phasianus (17). | 105. Meleagris (1). | |
| 105. Pucrasia (2). | ||
| 106. Chrysolophus (2). | ||
| 107. Crossoptilon (5). | ||
| 108. Lophophorus (1). | ||
| 109. Ithaginis (3). | ||
| 110. Caccabis (5). | ||
| 111. Francolinus (2). | ||
| 112. Perdix (2). | ||
| 113. Coturnix (2). | ||
| 114. Tetraophasis (2). | ||
| 115. Tetraogallus (5). | ||
| TETRAONIDæ (Grouse): | TETRAONIDæ: | TETRAONIDæ: |
| 116. Tetrastes (3). | 53. Lagopus (5.4). | 106. Colinus (3). |
| 117. Tetrao (5). | 54. Canachites (2) = Dendragapus (3) | 107. Oreortyx (2). |
| 108. Callipepla (3). | ||
| 109. Cyrtonyx (1). | ||
| 110. Bonasa (1). | ||
| 111. Tympanuchus (2). | ||
| 112. Pediocetes (1). | ||
| 113. Centrocercus (1). | ||
| TURNICIDæ (Fighting Quails): | ||
| 118. Turnix (3). |
By merely glancing over these lists we see at once how greatly the genera in either the first or third column exceeds those in the middle column. This tells us that the speciality of each region in relation to the other is very much greater than their similarity. In order, however, to bring out these differences and resemblances more clearly, and to show what they imply, the facts are summarised in the following table of the families, giving the distribution of their genera and species in the same order as in the preceding lists:—
FAMILIES OF NORTH TEMPERATE LAND BIRDS WITH THEIR DISTRIBUTION AS REGARDS THE PALæARCTIC AND NEARCTIC REGIONS.1
| Total Genera. | Families. | PALæARCTIC ONLY. | BOTH REGIONS. | NEARCTIC ONLY. | |||
| Genera. | Species. | Genera. | Species. | Genera. | Species. | ||
| 37 | Turdidæ | 30 | 185 | 3 | 0 | 4 | 18 |
| 1 | Cinclidæ | — | 4 | 1 | 0 | — | 1 |
| 1 | Accentoridæ | 1 | 11 | ||||
| 1 | Panuridæ | 1 | 1 | ||||
| 8 | Paridæ | 4 | 44 | 3 | 0 | 1 | 14 |
| 1 | Sittidæ | — | 8 | 1 | 0 | — | 4 |
| 2 | Certhiidæ | 1 | 3 | 1 | 0 | — | 1 |
| 9 | Troglodytidæ | — | 6 | 1 | 0 | 8 | 22 |
| 3 | Motacillidæ | 2 | 31 | 1 | 0 | — | 2 |
| 13 | Mniotiltidæ | 13 | 54 | ||||
| 1 | Vireonidæ | 1 | 12 | ||||
| 1 | Crateropodidæ? | 1 | 2 | ||||
| 1 | Oriolidæ | 1 | 2 | ||||
| 1 | Laniidæ | — | 22 | 1 | 0 | — | 2 |
| 2 | Ampelidæ | — | 1 | 1 | 1 | 1 | 2 |
| 4 | Muscicapidæ | 4 | 11 | ||||
| 7 | Hirundinidæ | 1 | 9 | 2 | 1 | 4 | 7 |
| 1 | Tanagridæ | 1 | 4 | ||||
| 41 | Fringillidæ | 15 | 110 | 8 | 5 | 18 | 79 |
| 8 | Icteridæ | 8 | 18 | ||||
| 8 | Alaudidæ | 7 | 26 | 1 | 1 | — | 1 |
| 4 | Sturnidæ | 4 | 8 | ||||
| 11 | Corvidæ | 4 | 30 | 4 | 2 | 3 | 14 |
| 9 | Tyrannidæ | 9 | 25 | ||||
| 4 | Cypselidæ | 1 | 8 | 1 | 0 | 2 | 4 |
| 4 | Caprimulgidæ | 1 | 4 | — | — | 3 | 4 |
| 4 | Trochilidæ | 4 | 11 | ||||
| 12 | Picidæ | 4 | 31 | 2 | 0 | 6 | 22 |
| 3 | Alcedinidæ | 2 | 5 | 1 | 0 | — | 2 |
| 1 | Coraciadæ | 1 | 2 | ||||
| 1 | Meropidæ | 1 | 3 | ||||
| 1 | Upupidæ | 1 | 1 | ||||
| 6 | Cuculidæ | 3 | 7 | — | — | 3 | 4 |
| 1 | Psittacidæ | 1 | 1 | ||||
| 1 | Strigidæ | — | 1 | 1 | 0 | — | 1 |
| 12 | Bubonidæ | 2 | 18 | 8 | 4 | 2 | 11 |
| 4 | Vulturidæ | 4 | 5 | ||||
| 3 | Cathartidæ | 3 | 3 | ||||
| 19 | Falconidæ | 4 | 51 | 10 | 5 | 5 | 26 |
| 6 | Columbidæ | 1 | 22 | 1 | 0 | 4 | 7 |
| 2 | Pteroclidæ | 2 | 8 | ||||
| 13 | Phasianidæ | 12 | 50 | — | — | 1 | 1 |
| 12 | Tetraonidæ | 2 | 14 | 2 | 1 | 8 | 20 |
| 1 | Turnicidæ | 1 | 3 | ||||
| 285 | (44 Families.) | 118 | 747 | 54 | 20 | 113 | 397 |
| Total in each region | 172 | 767 | 167 | 417 |
1 It must be noted that the numbers in the "Species" columns represent the species which are absent from the other region, not the number of species contained in the "genera" of the preceding column, Which are the genera absent from the other region.
The following statement, giving statistics derived from the preceding lists, will complete the information necessary for a comparative view of the relations of the Palæarctic and Nearctic Regions as regards their land-birds:—
The Palæarctic Region has 36 families of land-birds. Of these, 12 families are not in the Nearctic Region. These 12 families comprise 22 genera and 57 species.
The Nearctic Region has 32 families of land-birds. Of these, 8 families are not in the Palæarctic Region. These 8 families comprise 40 genera and 128 species.
The 118 Palæarctic genera which are not Nearctic comprise 472 species.
The 113 Nearctic genera which are not Palæarctic comprise 282 species.
The preceding lists and figures enable us to obtain a very complete view of the amount of difference that exists between the avifaunas of the Palæarctic and Nearctic Regions.
Considering first the Family groups, we find that in the one case one-third and in the other one-fourth of the families of the one region are not found in the other; and this is an amount of difference that does not occur between any other two regions which are conterminous with each other. The most striking relation of the Palæarctic and Nearctic Regions is, therefore, not their resemblance, but their dissimilarity.
Again, the 12 families which thus differentiate the Palæarctic Region from the Nearctic comprise 22 genera and 57 species; and among them we find such characteristic groups of the Eastern Hemisphere as the accentors, the flycatchers, the starlings, the vultures, and the sand-grouse,—families entirely absent from the whole Western Hemisphere.
The 8 families which differentiate the Nearctic Region from the Palæarctic are even more important, since they contain 40 genera and 128 species; and include such characteristic New World types as the Mniotiltidæ, the Icteridæ, and the Tyrannidæ, containing between them
nearly a hundred species. Now as there are only 417 species of land-birds in the whole Nearctic Region, we find that between one-third and one-fourth of the whole belong to families which are entirely foreign to the Palæarctic Region. It may be confidently asserted that none of the other regions can be so divided that the two parts shall show an amount of difference at all approaching to this. Yet it is proposed to unite these two regions because they are not sufficiently distinct. They are, however, very much more distinct than are the Ethiopian and Oriental Regions, though the former includes the isolated and peculiar Madagascar fauna. Less than one-twelfth of the Ethiopian families of land-birds are not Oriental, while only one-ninth of the Oriental families are not Ethiopian; showing that, by this test of the number of families which are not found in both regions, the Palæarctic and Nearctic are three times as distinct as are the Ethiopian and Oriental Regions.
If we now consider the genera which are characteristic of the one region as compared with the other, we shall find equally strong evidence of their diversity. In the Palæarctic Region we have 120 genera which are not Nearctic, out of a total of 174; so that a little more than two-thirds of the Palæarctic genera of land-birds are quite unknown in the Nearctic Region; and these genera contain 472 species out of a total of 767. In other words, out of every 5 land-birds in the Palæarctic Region, 3 belong to genera which are not Nearctic.
Looking at the same problem from the other side of the Atlantic, the results are even more striking. Out of a total of 167 genera of Nearctic land-birds no less than 113 are not Palæarctic, the proportion being almost exactly two-thirds. These 113 genera comprise 282 species out of a total of 417 species; so that again almost exactly two-thirds of the Nearctic land-birds belong to genera which are not Palæarctic. This is a larger proportion than in the case of the Palæarctic Region; and nothing can more forcibly bring before us the fundamental diversity of the two areas than the fact that, almost everywhere in the Nearctic Region, out of every three
birds we might meet with, two would be generically unknown to the student of the Palæarctic avifauna. There is probably no such amount of difference as this between any two adjacent regions, except perhaps between the Oriental and Australian, the latter admittedly the most isolated on the globe.
I have now shown, by a careful comparison of their mammalia and birds, that the Palæarctic and Nearctic Regions, instead of being so much alike that they should be united to form a single region, are really exceptionally distinct. They are certainly much more distinct than are the Oriental and Ethiopian Regions, and are probably quite as distinct as are any two conterminous regions.
I feel confident, therefore, that any naturalist who will study the materials I have here brought together in a form to admit of easy comparison, will arrive at the conclusion that the system of Zoological Regions established by Dr. Sclater cannot be improved by the union of two such fundamentally distinct areas as are those which he has termed the Palæarctic and Nearctic Regions. This conclusion entirely harmonises with the facts of plant-distribution as popularly described in the last two chapters.
CHAPTER XII
THE COLEOPTERA OF MADEIRA AS ILLUSTRATING THE ORIGIN OF INSULAR FAUNAS
THE Coleopterous fauna of the Atlantic islands is held by some writers to prove that the whole of these islands from the Azores to the Cape de Verdes, and even to St. Helena, are portions of a submerged continent, occupying a large part of the eastern Atlantic, and which was connected with, or formed an extension of, Southern Europe. And, further, that certain isolated affinities of African and American groups are believed to prove two distinct land-connections across the Atlantic, one between Brazil and Equatorial Africa, the other between Patagonia and South Africa.
The late Mr. Andrew Murray believed that, with the exception of the timber-borers, the presence of the same or closely allied species of beetles in discontiguous countries is a proof that there has been a former continuity of soil, because neither their powers of flight nor their vitality are sufficient to carry them over any considerable extent of sea. But in all these respects they are vastly inferior to mammals, reptiles and land-shells; while their generally small dimensions must offer facilities for distribution in many unexpected ways. Violent gales of wind, for example, will, we know, carry bodies of
1 This chapter formed a portion of the author's Presidential Address to the Entomolgical Society of London in January 1871, and is a reply to the arguments of the late Mr. Andrew Murray in a paper published by the Linnean Society in 1870, and to similar views held by many other writers, and still occasionally put forward.
greater specific gravity than beetles for many miles through the air; and storms and hurricanes are of such frequent occurrence, that they must have played a large part in stocking all uninhabited lands. Again, during great floods, whole forest trees are often carried out to sea, and hundreds of beetles may lurk in the crevices of their bark or even among their foliage, and, under favourable circumstances, be drifted a long way in safety. Even matted rafts covered with soil and bearing living vegetation are occasionally floated out to sea by tropical rivers and may be drifted along for weeks, and ultimately convey scores of insects to far distant lands. A large number of beetles are exceedingly tenacious of life. Immersion in strong spirits for twelve hours will not always kill them, nor will water if much below the boiling point; so that it is not improbable that some considerable proportion would be found to survive immersion in sea-water for several days. Many facts have not been recorded as to the passage of beetles over wide tracts of ocean, but some of them are sufficiently remarkable. Darwin captured a Colymbetes forty-five miles from land north of the Rio de la Plata; and at seventeen miles off Cape Corrientes he caught in a net a number of live beetles of the genera Colymbetes, Hydroporus, Hydrobius, Notaphus, Cynucus, Adimonia, and Scarabæus. A Calosoma also flew on board the Beagle when ten miles from the shore of South America, and the Calosoma sycophanta is believed occasionally to cross the English Channel. Sir Charles Lyell also states that exotic beetles are sometimes thrown on our shores, which revive after being long drenched in salt water. In the case of other insects, we have more positive proof of their passage over wide spaces of ocean. A whole swarm of locusts has been known to pass over Madeira from Africa, a distance of more than 300 miles; while Darwin himself captured a locust at sea 370 miles from land. Two individuals of the Sphinx atropos flew on board the Hotspur East Indiaman in 1866, during an easterly gale, at a point 260 miles from the coast of Portugal, and were exhibited at a meeting of the Zoological Society. In his work on the Natural History of the Azores, just
published, Mr. Godman relates that a white butterfly flew on board a whaler coming from the south, at about 400 miles from the Azores: it was caught by the captain and placed in a drawer, where it laid several eggs.1 Such cases as these having been already recorded, we may be sure that migrations to much greater distances are constantly occurring, since we can hardly suppose the extreme cases to be those which have first been observed. We have therefore every reason to believe that, under favourable conditions, almost any winged insect could traverse equal distances. These considerations would lead us to the conclusion that a partial identity of species may exist in the beetles of two countries separated by some hundreds of miles of sea, without in any way necessitating the former existence of a continuity of land between them. In the case of the Atlantic islands, therefore, I see no reason to believe that they owe their Coleoptera to a land-connection with the continent, more especially when there is such strong evidence against that view in the total absence of all mammals and reptiles. Can we believe that the forests of Madeira would be without a single native rodent, or even a frog, if they owed their rich coleopterous and molluscous faunas to land-connection with Europe?
The exhaustive researches of Mr. Wollaston in these islands will, I believe, furnish, in the single order of Coleoptera, ample materials for the elucidation of this very interesting question. Although the Insecta Maderensia has now been published more than forty-six years, the vast store of facts which it contains bearing on the question of geographical distribution, and especially on that of insular faunas, has never been fully appreciated; and as neither Mr. Murray nor later writers have grappled with these facts as a whole, or attempted to show how they are compatible with their theory, I think it advisable to give a somewhat detailed analysis of them, pointing out what I conceive to be their true bearing on the problem of the mode of distribution of beetles and the origin of insular faunas. My interpretation of the evidence may be erro-
1 Other cases are given in my Darwinism, p. 359.
neous, but the facts themselves must be of value. The outline map here given will show the relation of the islands to the other Atlantic groups and to the adjacent continents.
I propose to confine myself mainly to evidence furnished by the Coleoptera of the Madeiran group, because, being
FIG. 51.—OUTLINE MAP OF THE AZORES.
| NOTE.— | The light tint shows where the sea is less than 1,000 fathoms deep. |
| The dark tint „ „ „ more than 1,000 fathoms deep. The figures show depths in fathoms. |
separated from the mainland by a much wider extent of ocean than either the Canaries or Cape de Verdes, they offer a much more satisfactory test of the opposing theories. It is an advantage also that the materials are by far the most complete; and in the Insecta Maderensia Mr. Wollaston has given some details of importance which are
wanting in the Coleoptera Atlantidum and in the Coleoptera Hesperidum.
The most novel and striking facts brought out by Mr. Wollaston's researches in Madeira are, as is well known,— 1st. The affinity with the Mediterranean fauna;—2nd. The total absence of certain large divisions of Coleoptera abundant in that fauna;—3rd. The number of new and peculiar species and of new and anomalous genera;—and 4th. The unexampled preponderance of apterous (wingless) species. If we accept, as all naturalists now do, the theory of slow change of forms by natural causes, we may take the first and third of these facts as proving that the origin of the Madeiran fauna is of very ancient date. Let us see therefore how the second and the fourth set of facts bear upon the mode of its origin, whether by a land-connection with Europe or by transmission across the sea. It will be convenient to take first the facts presented by the wingless or winged condition of the species.
This striking peculiarity consists, either in species being apterous in Madeira which are winged elsewhere, or in genera which are usually winged consisting of only apterous species in Madeira, or lastly in the presence of endemic apterous genera, some of which have winged allies while others belong to groups which are wholly without wings. Such phenomena undoubtedly show that there is something in Madeira which tends to abort wings; and Mr. Wollaston was himself the first to suggest that it was connected with exposure to a stormy atmosphere. His further observation, that many of the winged species had wings more developed than usual, enabled Mr. Darwin to hit upon that beautiful explanation of the facts which commends itself to all who believe in the theory of Natural Selection; while Mr. Wollaston himself admits it as fully accounting, teleologically, for the phenomena. That explanation briefly is, that the act of flying exposes insects to be blown out to sea and destroyed; those which flew least therefore lived longest, and by this process the race became apterous. With species to whom flight was a necessity, on the other hand, the strongest winged lived longest,
and thus their wings became more and more developed in each succeeding generation.
Now this view of the case enables us at once to explain some of the most striking gaps in the Madeiran coleopterous fauna. The Cicindelidæ, for instance, are entirely absent; and almost all the European species are winged insects of somewhat feeble flight, yet to whom flight is necessary. We can readily understand that such insects would be easily exterminated if they arrived singly or in small numbers; though it is not so easy to understand why, in a forest-clad island, some of the sylvan species should not have found a home had the land ever been connected with a continent where they abound. Their total absence is, therefore, decidedly unfavourable to the theory of a land-connection with Europe. To the Melolonthidæ and Cetoniidæ, as well as the Eumolpidæ and Galerucidæ, which are all wanting, the same argument will apply; and also to the Elateridæ and Buprestidæ, which are represented each by one minute species. But if Madeira is the remains of a continent once continuous with the south of Europe and deriving its fauna from such continuity, how are we to explain the absence of extensive genera very abundant in South Europe, and, from their being apterous, specially adapted to the peculiarities of Madeira? Such are Carabus, Lampyris, Pimelia, Akis, and many others. But these facts are all consistent with the theory of introduction across the sea. Apterous groups, however abundant on the continent, should, as a rule, be absent; and I find that almost all the European apterous genera are wanting, and among the few exceptions there are some whose presence is easily explained and really prove the rule. We must remember, however, that the apterous condition, except in those cases where it is characteristic of an extensive group, is one of little stability or importtance. There are species which are sometimes apterous and sometimes winged, and we may therefore be sure, that if any advantage was to be derived by either condition over the other, natural selection would very rapidly render it constant by the repeated survival of the favoured individuals. This is illustrated by the fact that we have
winged and apterous species in the same genus, as well as winged and apterous genera in the same family. The Coleopterous Order being essentially winged, and the vast majority of its members being capable of flight, it is a presumption, if not almost a certainty, that all apterous varieties, species, or groups, have been derived from winged ancestors,—comparatively recently in the case of the former, and at a more remote epoch as the character becomes more constant and attached to groups of higher classificational value.
Taking these principles as our guide, let us examine more closely the facts presented by the Madeiran Coleoptera, and their bearing on the rival theories as to their mode of introduction.
There are a large number of European beetles belonging to very varied genera and families which are apterous, and a large proportion of these inhabit the South of Europe and North Africa. Now, on the theory of land-connection, there should be no marked absence of these groups; on the contrary, apterous forms being especially adapted to Madeira, we should expect them to predominate. But, on the opposing theory of transmission across the sea, we should expect them to be wholly absent, or, if there are any exceptions, we should expect to be able to detect some special circumstances which might favour their transmission. A careful examination of Lacordaire's Genera, and of some works on European Coleoptera, has furnished me with the following list of genera which are wholly apterous, and which abound in South Europe and North Africa.
Carabus possesses about 80 species in these regions; but is wholly absent from Madeira.
Thorictus has 10 South European species, and one representative in Madeira, which is an ants'-nest species.
Rhizotrogus (Melolonthidæ), 27 species in Sicily and Algeria, the very countries to which the Madeiran fauna is traced, yet it is wholly absent.
Lampyris, Drilus and Troglops (Malacoderms), of which the females are apterous, possess 27 South European and North African species; none in Madeira.
Otiorhynchus, Brachycerus, and twenty other genera of Curculionidæ, comprising more than 300 South European and North African species, are absent from Madeira, with two exceptions. One is the Trachyphlœus scaber, a widely-spread European insect often found in ants' nests; and this, with the case of the Thorictus, renders it probable that ants'-nest species have some unusual means of distribution, which are by no means difficult to conceive. The other exception is that of the genus Acalles, which has a number of Madeiran species, all peculiar, and is very abundant in all the Atlantic islands. Now we have first to remark that Acalles is an isolated form, but is allied to Cryptorhynchus, which is often amply winged; so that we may easily suppose that its introduction to Madeira took place before it became completely apterous in Europe. In the second place, we have the fact that many of the species are confined to peculiar herbaceous and shrubby plants, in the stems of which they undergo their transformations, and which habit would afford facilities for their occasional transmission in the egg or pupa state across a considerable width of ocean, while a fragment of dry stem containing egg or larva might possibly be carried some hundred miles or more by a hurricane. Such suppositions would not be admissible to account for numerous cases of transmission, but, as will be seen, this is almost the only example of a genus of large-sized apterous European beetles occurring in Madeira.
Pimelia, Tentyria, Blaps, and eighteen other genera of Heteromera, comprising about 550 species of South Europe and North Africa, are totally absent from Madeira, with the following interesting exceptions:—two common species of Blaps, which are admitted to have been introduced by human agency, and three species of Meloe, two of which are European and one peculiar. The means by which the apterous, sluggish and bulky Meloes were introduced is sufficiently clear, when we remember that the minute active larvæ attach themselves to bees, insects of exceedingly powerful flight, and more likely than perhaps any others to pass safely across 300 miles of ocean. That the solitary exception to the absence of wholly apterous
VOL. I. S
genera of European Heteromera from Madeira should be the genus Meloe, is, therefore, one of those critical facts which almost demonstrate that it is not to land-continuity with the continent that the island owes its insect fauna.
Timarcha. This, the only important apterous genus of Chrysomelidæ, is especially abundant in Spain and Algeria, and possesses forty-four South European and North African species; yet it is unknown in Madeira.
The occurrence of two isolated European species of characteristic Atlantic apterous genera—Tarphius and Hegeter—may seem to favour the opposite theory. The Tarphius gibbulus occurs in Sicily, and is the only European species of the genus, of which about forty inhabit the Atlantic islands. It is most nearly allied to the smallest of the Madeiran species, T. Lowei, which is abundant among lichen on weather-beaten rocks and even ascends in the forest regions to the highest branches of the trees. These habits, with its minute size, are all in favour of this species, or some ancestral allied form, having been carried across by the winds or waves, thus transferring to Europe one of the peculiar types elaborated in the Atlantic isles. The Hegeter tristis is an analogous case, this species of an otherwise exclusively Atlantic genus having occurred on the opposite coast of Africa. These instances will furnish a reply to one of Mr. Murray's difficulties,—that all the migration has been in one direction, from Europe to Madeira, never from Madeira to the continent,—a difficulty, it may be remarked, which is wholly founded on an unproved and unprovable assumption; for how can it be determined that, in the case of Acalles for example, the genus had not been first developed in the Atlantic islands and then transferred to Europe, instead of the reverse? It is always assumed to have been the other way, but I am not aware that any proof can be obtained that it was so, and it is inadmissible to take this unproved assumption, and base an argument upon it as if it were an established fact.
We will next consider the facts presented by the distribution of those species of Coleoptera which range from Madeira to Europe, or to any of the other Atlantic islands.
If their distribution has been effected by land-continuity, we should expect that the proportion of winged and apterous species that extend their range beyond the island, should not be very strikingly different from the proportion that is found on the island. We do not find, for example, that the proportion of the wingless Carabi that have reached our own country from the Continent by former land-connection is very different from that of the winged Cicindelæ.
Now, leaving out altogether those species which have certainly been introduced by man, and grouping the remainder for convenience in six divisions, we find that the Madeiran Coleoptera, which are not peculiar to it, may be classed as follows:—
31 species of Carabidæ, of which 26 are winged, 5 apterous. The whole fauna, however, presents the very different proportion of 38 winged, 43 apterous.
93 species of the families from the Hydradephaga to the Tomicidæ inclusive, of which 90 are winged, 3 apterous. Total fauna: 220 winged, 27 apterous.
28 species of Curculionidæ, of which 26 are winged, 2 apterous. Total fauna: 35 winged, 74 apterous.
15 species of Longicornia and Phytophaga, of which 15 are winged, none apterous. Total fauna; 48 winged, 1 apterous.
20 species of Heteromera, of which 16 are winged, 4 apterous. Total fauna: 28 winged, 27 apterous.
76 species of Staphylinidæ, of which all are winged, none apterous. Total fauna: 109 winged, 6 apterous.
The totals are, for the wide-ranging species, 249 winged, 14 apterous = 263; for the whole fauna, 478 winged, 178 apterous = 656.
It thus appears that, in every case, an immensely smaller proportion of apterous than of winged species are widely distributed. If we take the totals we find that while about two-fifths of the whole number of species range to other countries, only about one- thirteenth of the apterous species do the same, although among the strictly endemic species there are 160 apterous to only 110 winged! We can
S 2
hardly impute such a constant and overwhelming preponderance to the fact that apterous insects have less facilities for extending their range, when we know that nearly every apterous genus possesses species of almost universal European distribution. I may here recall the fact, that of the above-mentioned 14 apterous species which range out of Madeira, two are Meloes and two ants'-nest beetles, whose presence we have already sufficiently accounted for. It may no doubt be said that much of the difference here shown is due to the fact that the peculiar Madeiran species have had time to become apterous, while the species common to other countries have not yet had time to lose their wings; but this argument, although a valid explanation of some portion of the facts, if we admit that many of the latter have been recently introduced by natural causes, cannot be used by those who maintain a former land-connection as the sole origin of the fauna; for on that theory all the species now inhabiting the island (and not introduced by man) must date back to the same remote period, and have had equal time in which to be modified.
Let us now consider what are the special relations of the apterous Madeiran species as throwing light upon their possible or probable mode of introduction.
We have three species which Mr. Wollaston himself states to be usually winged elsewhere, but which are apterous in Madeira. These are Metabletus obscuroguttatus, Calathus fuscus and Bradycellus fulvus. I am inclined to believe that there are a few others which will come under this category, but it is very difficult to get information as to the winged or apterous character of particular species. These insects, however, have evidently become apterous since their introduction into Madeira. We have therefore no difficulty in accounting for their introduction, and, as no other change in their external characters has been effected, we may suppose it to have been comparatively recent.
Next we have those genera which, though apterous in Madeira, are wholly or partially winged elsewhere. These comprise a large number of species, and are twenty-two
in number, as follows:—CARABIDæ: Cymindis, Dromius, Metabletus, Scarites, Apotomus, Loricera, Leistus, Calathus, Olisthopus, Argutor, Cratognathus, Bradycellus, Trechus. PHILHYDRIDA: Hydrobius. BYRRHIDæ: Syncalypta. CURCULIONIDæ: Phlœophagus, Tychius, Smicronyx. HETEROMERA: Phaleria, Helops. STAPHYLINIDæ: Homalota (1 sp.), Othius. Here we are carried back to a remoter epoch for the introduction of the winged ancestors of the Madeiran species, since not only have the wings become aborted, but the insects themselves have become modified into distinct and often very well-marked species.
The next category consists of apterous genera which are peculiar to Madeira and the other Atlantic islands, but which are allied to winged groups, as follows:—
Elliptosoma.—Closely allied to Loricera, winged.
Eurygnathus.—An abnormal form of Licinides, most of which are winged.
Zargus.—An abnormal form of Chlæniides, winged.
Thalassophilus.—Allied to Trechus, winged.
Tarphius.—Belonging to the Colydiidæ, most of which, Mr. Pascoe informs me, have wings.
Coptostethus.—Allied to Cryptohypnus, winged.
Caulophilus.—Allied to Phlœophagus, winged.
Lipommata, Mesoxenus, Caulotrupis. — Anomalous genera of Cossonides, which are often winged.
Acalles, Torneuma.—Aberrant genera of Cryptorhynchides, most of which are winged.
Echinosoma.—Doubtful affinities.
Atlantis, Cyphoscelis, Laparocerus (Laparocerides).—A very isolated group.
Anemophilus, Scoliocerus.—Allied to Trachyphlœides, some of which are winged.
Lichenophagus.—Allied to Cænopsis and Omias, some of which are winged.
Xenorchestes.—Allied to Choragus, winged.
Ellipsodes.—Closely allied to Crypticus, some of which are winged.
Hadrus.—Belongs to an apterous group of Opatrides, many of which are winged.
Macrostethus.—Belongs to Cœlometopides, all of which
are apterous, but comes next to the Tenebrionides vrais, of Lacordaire, which are mostly winged.
Xenomma.—Belongs to the Aleocharides, which are winged.
Mecognathus.—Allied to Sunius, winged.
Metopsia.—Allied to Phlœobium, winged.
Here we have indications of an introduction of forms at a still more remote epoch. In many cases the modifications of structure have been so great as to produce distinct generic forms, while these remain still allied to winged European genera. In other cases, however, the modifications are still greater, and the affinities are with groups which in Europe are wholly apterous. Such cases as Hadrus and Macrostethus, which belong to small groups of wholly apterous genera, are difficulties on the theory of transmission over the sea. But two considerations render this difficulty less real than apparent. They all carry us back to a very remote epoch; and, knowing what we do of the instability of the apterous condition, we may fairly conclude that the groups in question were, at that time, in a partially winged state. At or near this same remote epoch, the Madeiran group, as indicated by the submarine bank now connecting the several islands, probably formed one more extensive island, and the distance of ocean to be traversed would then have been considerably less than it is now.
If the various groups of facts which I have here set forth, respecting the distribution of apterous and winged species and genera, are fairly considered as a whole, I think they will be seen to be quite inconsistent with the theory of that distribution having been effected by a former land-connection with Europe; and, considering that we are necessarily ignorant of many of the ways by which organisms are transmitted across ocean barriers, such transmission seems to be indicated in the case of the Madeiran Coleoptera, not by means of drift wood and ocean currents, which Mr. Murray thinks must be the most efficient means of transport, but by some mode in which their wings are called into play, which can only be by a passage through the air when assisted by gales and hurricanes.
There is one other group of islands which seems well adapted to offer a crucial test of the correctness of the theory of land-connection. The Azores are more than twice as far from Europe as the Madeiras, and, what is of still more importance, they are cut off from it as well as from the Madeiras by a broad belt of ocean of the enormous depth of nearly 15,000 feet. We may feel pretty confident, therefore, that if both groups have once been united to the continent, the separation of the Azores is by far the more ancient event; and any theory which requires the Azores to be the most recently separated must be strongly supported by independent evidence to render such an improbable supposition acceptable. If the Azores date the origin of their insect population from a remote epoch when they were connected with Europe, we should expect to find that almost all the species have since become modified, and that these islands would offer us a larger proportion of highly specialised and ultra-indigenous forms than Madeira itself. The exact contrary, however, is the fact, for out of more than 200 species only about sixteen are peculiar. Taking the geodephagous group, the species of which, both Mr. Murray and Mr. Wollaston believe, are least liable to be introduced by man, we find that two only are peculiar, while sixteen are European. The Rhynchophora only equal the Geodephaga in number of species, and seven of these are peculiar. Leaving out a large number of species which have, there is little doubt, been introduced through human agency, there remain more than 100 species identical with those of Europe and the Atlantic islands, while only fourteen are peculiar. These facts imply that the insects, as a whole, have been brought to the islands through natural causes, and that the process is probably still going on. On looking to Physical Maps for information, however, a difficulty appears; for the ocean currents, as well as the prevalent regular winds, are all from the westward, while only four of the beetles are American, and these being all wood-borers have no doubt been brought by the Gulf-stream or in some cases have been introduced by man. Fortunately, however, we have a means of getting over this difficulty; for Mr. F. Du Cane
Godman, who has given us the most recent and accurate information on the natural history of these islands, informs us (in his paper on the Birds of the Azores in the Ibis for 1866) that the stormy atmosphere, to which we have seen that Madeira owes so many of its peculiarities, is still more marked a feature of the Azores, where violent storms from all points of the compass are frequent, and annually bring to their shores numbers of European birds. As a natural result of this constant influx, the birds of the islands are, all but two, of European species; and, what is very important, they decrease in numbers from the eastern to the western islands of the group. This is just what we should expect if they are stragglers from the eastern continent; but if they are the descendants of those which inhabited the country before its dismemberment, there would be no meaning in such a diminution. Now we can hardly doubt that these same storms also bring Coleoptera and other insects to the Azores, though it may be more rarely and in smaller numbers than in the case of birds; and the large proportion of European species will then be very intelligible. The same explanation is suggested by the proportions of the most important groups, for while (after deducting all those species believed to have been introduced by man) the Geodephaga and Brachelytra are by far the most numerous, the Rhynchophora and the Heteromera are exceedingly few, a distribution which corresponds with their respective powers of flight. It is also a very important fact that only four non-introduced species can be traced to an American origin, while more than a hundred are European; since it shows of how little importance are ocean currents as a means of conveying insects over a wide extent of sea; whereas the great mass of the non-introduced species have evidently passed through the air, aided by their powers of flight, for a distance of about a thousand miles from Europe. The Azorean Elateridæ form a curious feature of its fauna, considering that the whole family is almost absent from Madeira and the Canaries. Of the six species two are European (one specially Portuguese), so that they may have been introduced with living plants. Two are common
South American species, probably introduced in the floating timber, though they may also have come with living plants, which are often brought from Bahia. Two species, however, are peculiar, and one is closely allied to a Brazilian species, so that it must have been introduced by natural agencies before the settlement of the island; the other is of a genus confined to Madagascar.
Now it is a suggestive fact that the Mozambique current, bending round the Cape of Good Hope to the Equator, is one of the sources of the Gulf-stream; so that it is not impossible that a tree, carried down by a flooded river on the west coast of Madagascar, might ultimately reach the Azores. That it should convey living larvæ or pupæ of Elaters may also not be impossible; and if such a log reached the Azores but once in ten thousand years, and but one log in a thousand should convey living Elaters, we should still, if the calculations of geologists have any approximate value whatever, be far within the epoch of existing genera, and even of most existing species. A relation so isolated and extraordinary as that between a single insect of the Azores and those of Madagascar, may well be due to a concurrence of events as rare and improbable as this seems to be.
The Azores, and in a less degree the Madeiras, appear to me to teach us this important lesson in the laws of distribution of birds and insects,—that it has been determined neither by the direction of ocean currents nor by that of the most prevalent winds, but almost wholly by more exceptional causes such as storms and hurricanes, which still continue to bring immigrants from the nearest lands.
Mr. Murray's argument for a land-connection between the various Atlantic islands, from the Azores to the Cape de Verdes, and even to St. Helena, has perhaps more to be said for it; but I do not think that the facts require anything beyond the extension of each group into one or more considerable islands. Such an extension is perhaps indicated by the comparatively elevated submarine bank on which each group stands; and it is evident that more extended land-surfaces would not only bring the groups nearer to each other, but, by offering a much greater length
of opposing coasts, would greatly facilitate the migration and accidental transmission of individuals.
Another point of great importance to which attention is called in Mr. Murray's essay, is the long-persisting identity of form which seems to be a characteristic of insects, and which is thought to allow ample time for those revolutions in geography to which he so constantly appeals. But this antiquity and persistence of insect-forms will have allowed equal time for the action of a most powerful agent of distribution, which is too hastily dismissed. I allude to those changes of climate, which within a period so recent as the Miocene, have at one time clothed the now inhospitable regions of North Greenland, Spitzbergen and other Arctic lands, with forests and evergreens and flowering shrubs, and at another have covered the Northern United States and Central Europe with a mantle of ice like that which at present envelopes Greenland. These vast climatic changes must have afforded ample facilities for insect migrations,—between the eastern and western hemispheres, when the arctic regions were inhabited by a temperate flora and fauna,—between the northern and the southern, when the animals and plants of either hemisphere were driven towards the Equator by the glaciation of their native regions, and when a portion would cross that barrier, either along the elevated lands or by transmission over narrow seas. This cause is admitted by our best botanists to be amply sufficient to account for the presence of European genera and species of plants on the Andes, in Chili, Patagonia and Terra del Fuego, in New Zealand, and in the Australian Alps. The relations of South America, Australia, and other southern lands to each other, are still more marked, and probably more deep-seated, and seem to imply either a greater extension or the existence of intermediate lands at some former period, but not an actual continuity with one another.
I believe that the curious and suggestive facts which I have disinterred from that bulky and little-read volume, the Insecta Maderensia, may be of some use to students of the geographical distribution of animals.
CHAPTER XIII
EVOLUTION AND THE DISTRIBUTION OF ANIMALS1
IN the Nineteenth Century of December 1878, Dr. P. L. Sclater called attention to the subject of the geographical distribution of animals in its bearing on the theory of evolution, and gave numerous special cases in which the actual distribution of particular species and groups is very difficult to explain on that theory without making assumptions which, in his opinion, the evidence at our disposal does not warrant. Difficulties of this nature are so numerous, and many of them seem to him so weighty, that in order to explain them, he is led to question, what is almost an axiom with evolutionists, that identity of structure is, without exception, an indication of descent from a common parent. Similar doubts, though not stated in exactly the same terms, were felt by the late Professor Mivart2; and it therefore becomes a matter of interest to examine a little more closely into the alleged difficulties, in order to see whether they are not really explicable on the principle of descent with modification, only calling to our aid such general assumptions as are fully warranted by what we actually know of the migrations and extinctions of living things, and of the past changes in the physical condition of the earth and its inhabitants.
As Dr. Sclater's article gives an excellent summary of the nature and meaning of zoological distribution, and of
1 This chapter first appeared in the Nineteenth Century of February 1879, under the title "Animals and their Native Countries."
2 Genesis of Species, Chap. III.
the main general conclusions arrived at by naturalists, our purpose will be best attained by proceeding at once to consider his special cases of difficulty; and in doing so we shall have occasion to discuss, as fully as may be required, the general principles and particular illustrations needed to elucidate them.
We have first the case of the Little Blue Magpie of Spain (Cyanopica Cooki), which has a very close ally in the extreme parts of Eastern Asia and Japan (C. cyaneus), while there is nothing closely allied to these in all the intervening regions or in any other part of the world. This is said to be an infringement of the canon as to the continuity of specific areas, and as such to require explanation. Before proceeding further, it will be well to inquire into the value of this canon of continuity, and whether it is so clear and well established that infringements of it are altogether exceptions to the usual course of nature. So far from this being the case, I believe it will be found that, between the complete continuity of the area occupied by a species or a genus and such wide discontinuity as occurs in the present example, there is every possible gradation; and further, that the instances of discontinuity are very numerous, while those of complete continuity are far less generally the rule than appears at first sight.
In order to understand the bearing of this class of phenomena on the theory of derivation by modification from an allied form, let me briefly indicate the probable course of a genus of animals from its birth or origin to its final extinction.
Genera are groups of species which agree among themselves, and differ from all other groups in the same family or order, by the possession of some structural peculiarities. We must therefore suppose a genus to have had its origin in some variation of structure which was useful to its possessors—such as a modification of the bill, feet, or wings of a bird, or of the teeth, claws, or horns of a mammal. According to the theory of natural selection, the possessors of such a useful peculiarity would increase at the expense of their close allies who did not possess it,
and would soon form a distinct group of individuals breeding together and constituting a species—the first species of the new genus. This species having in time supplanted the parent species, and being better adapted than it for the battle of life, would almost certainly cover a wider area, and thus come into competition with several of the allied species of the old genus, some of which it would also probably supplant, and occupy the areas they formerly occupied. But as the old genus had been modified into distinct species (differing, perhaps, slightly in colour or habits in accordance with the varying physical conditions), so the now widespread species of the new genus would vary, and become modified in a somewhat analogous manner, forming a genus consisting of several species. Now, if the generic form thus produced was one of great inherent vigour and adaptability, and if the peculiarity of structure it possessed was of considerable importance, it would become what Mr. Darwin terms a dominant group: that is, it would spread widely over the earth under various modified forms suited to the various conditions it became subject to. At last it would reach its maximum of development, and cease to spread further, either owing to its inability to adapt itself to further changes of climate, &c., or, what is more likely, from its coming into competition with other dominant groups which had in like manner spread from some other centres.
Now, during all this time, which may be termed the period of growth of the genus, its area will have been almost necessarily continuous, and the areas occupied by its several species (also continuous) will probably overlap each other. But now commences its period of decay. Other groups of the same or allied families have given rise to varieties which have also become dominant species and genera, which, under the somewhat changed physical conditions that in time have come about, beat it in the battle of life, and force it to retire step by step from the vast area it had overrun. First one species and then another will dwindle away and become finally extinct, and by so doing will necessarily leave gaps in its area of distribution. This process going steadily on, the time will
at last come when two or three species only will remain, most likely in widely separated parts of its former area; their position being determined either by the competition being there somewhat less severe, or by some speciality of conditions which are exceptionally favourable to the dying-out group. Then one and then another of these species will die out, and the once extensive genus will only be represented by a single species inhabiting a very restricted locality. This will become rarer and rarer, the necessary preliminary to that final extinction which we know to be the fate, sooner or later, of every group of living things.
Most working naturalists (and none better than Dr. Sclater) are acquainted with genera whose distribution will illustrate all the successive phases of this hypothetical history; while palæontology furnishes us with some actual examples of the progress of a group from its rise to its decay, though, owing to the extreme imperfection of the geological record (and its total absence for important epochs in many parts of the globe), we can never trace the complete history of such a group. A little consideration will show us, however, why it is that continuity of generic and specific areas appears to be the rule, discontinuity the exception, although the reverse may really be the case. There can be no doubt that the development of an extensive genus is a slow process, while its decay and final extinction need not be slow, and may conceivably be extremely rapid. Geological and geographical changes may be long in preparation, but finally very abrupt. Land may sink a thousand feet without producing any very important effect except diminution of area, but the next hundred feet of depression may cut it off from a continent, and may alter the direction of ocean currents, thus producing a greater organic and physical change than had been brought about by the previous subsidence occupying ten times as long. Again, such a change as that which admitted the highly organised Miocene mammalia of Europe into Tropical and South Africa must have led at once to the extermination of many of the indigenous species, and have restricted the
area of many more. It is also important to remember that the dominant or growing species and genera, which are those having continuous areas, will be necessarily more prominent, more numerous in species and individuals, and therefore far better known; while those in process of extinction, and for that very reason having discontinuous areas, will be less numerous, far less common, and in fact often very rare, and therefore much less known. In many cases, too, it will happen that the discontinuity is not great as regards distance, and it will then not be noticed, or will be imputed to want of knowledge, although it may be quite as real as when half a continent lies between the two species.
It appears, therefore, that the discontinuity of many genera and higher groups, so far from being difficult of explanation, is really one of the inevitable results of the process of extinction which is always going on. The peculiarity of the particular case we are considering is that it is somewhat extreme in the fact of two species only being left, occupying limited areas situated at the opposite extremities of the immense Palæarctic region. But this is not very extraordinary, because there are in Western Europe and Japan a number of pairs of closely allied species whose extinction in the intervening areas would lead to an exactly similar phenomenon to that we are considering. Such are the European and Japanese jays, bullfinches, goldcrest warblers, and wrens, all of which are closely allied to each other, while they are separated by a wide area in Central Asia often occupied by species which differ considerably from both. Should either of these groups die out, we might expect that the species inhabiting the comparatively desert and inhospitable regions of Central Asia would succumb first, while those living in the milder and more equable climates of Western Europe and Japan would probably linger on, the last of their race. It is very interesting to note that in most cases of such widely separated but closely allied species or groups there is a decided similarity in the general physical conditions of the countries they inhabit. The ally of the Spanish blue
magpie is found in North-eastern Asia from Shanghai to Pekin and the Amoor, as well as in Japan; and these countries reproduce the hot summers and the cold winters, the rugged mountains and the sheltered valleys of Spain, while both areas are subject to the influence of the vicinity of the ocean in an almost equal degree.
Before going further we must guard against a misconception as to the progressive rise and decay of species, genera, and higher groups. It is not maintained that this will always take place uninterruptedly or continuously. On the contrary, it is certain that the decay of a group may run its course for a time, and then, owing to changed conditions, may be checked, and even be changed into a new growth and development. Hence arise those isolated groups, which yet, by their abundance in species and the considerable area they occupy, show that they are in a flourishing condition—of which the tanagers in South America (Tanagridæ), the broadbills (Eurylæmidæ) of Asia, and the colies of Africa (Coliidæ), may serve as examples. When an extensive group is in process of extinction, it may become broken up into many isolated portions, sometimes in juxtaposition to each other, sometimes separated in remote parts of the globe. Changes of conditions, whether physical or organic, may favour first one, then another, of these portions, or the same portion may be subject to alternate phases of progression and decay several times repeated. How can we wonder that the final result of such complex processes, whose general nature we can understand but whose details it is impossible for us to trace, should often lead to anomalies in geographical distribution? And when we consider that these processes have been often intensified and further complicated by geographical mutations, and by those forced migrations induced by the climatal changes which culminated in the glacial epoch, the wonder rather is that we can account for so much, than that there should be matters of detail which we cannot explain.
It is clear, then, that the case of the Little Blue Magpie of Spain and its close ally in Eastern Asia is simply an
example of a dying-out group, of which two allied species remain isolated in remote countries favourable to their existence; and further, that the supposition of their common ancestors having once occupied the intervening region, so far from being unwarranted, is supported by the analogy of several other groups of birds in the same area.
Dr. Sclater's next two cases may be sufficiently explained by the application of the general considerations and examples already adduced. We have two allied species of Oxyrhamphus isolated in South-East Brazil and Central America—countries, it may be remarked, about equally removed from the equator and enjoying very similar climates; while in the cuckoos of the genus Neomorpha we have a similar phenomenon in nearly the same two areas, with the addition of three species in the intervening districts which are not closely allied to the two others.
We have here really only the same class of facts as occur plentifully in the Palæarctic region, in which the species of the eastern and western extremes are often alike, while those that intervene are more diverse. We can, without much difficulty, refer this latter peculiarity to diversity or similarity of climate and physical conditions, while in the Neotropical region it is more probable that a diversity of organic conditions may have been the agent at work. There is some reason to believe that the great plateau of Guiana long formed an island, and that this isolation led to the development of several peculiar forms, which have in some cases spread into Upper Amazonia. A range of plateaux and hills, on the other hand, connects Brazil with the Andes, and has thus kept up a greater zoological continuity with Central America than the intervening area of Guiana has been able to do.
Dr. Sclater's third problem, that of Pitta angolensis, is very interesting, and will afford us an opportunity of discussing some of the most curious phenomena of distribution, and of bringing forward some considerations which I believe will go far towards the removal of most of the difficulties they present. The case is that of a rather
VOL. I. T
extensive and very beautiful group of birds, though not very numerous in species, ranging from the Himalayas to Eastern Australia, but having one species isolated in Western Africa. Dr. Sclater has himself mentioned a few parallel cases, but there are others equally interesting, a consideration of which may aid us in our attempted explanation. We have first the gorilla and chimpanzee, corresponding to the orangs of Borneo and Sumatra; and among the quadrumana the genus Cercocebus allied to the Eastern macaques is wholly West African, while Colobus, closely allied to the Asiatic Semnopithecus, is found in Abyssinia as well as in West Africa. Among birds we have Alethe, an African genus of babblers, and Pholidornis, one of the Dicæidæ, allied to Asiatic forms; while the genus of crested hornbills, Berenicornis, has one species in Sumatra and the only other in West Africa. The fruit-thrushes of the genus Criniger, so abundant in Asia and Malaya, are also found in West and South Africa, while the beautiful parroquets of the genus Palæornis inhabit West Africa and Abyssinia as well as India and the larger Malay Islands. Among reptiles and amphibia we have three families which follow the same rule. Lizards of the family Acontiadæ are confined to the Moluccas, Ceylon, and West and South Africa; toads of the family Engystomidæ have nearly the same range, but are more widely spread in Asia; while snakes of the family Homalopsidæ are abundant in tropical Asia and America, and are even found in Europe, while in Africa they are confined to the western districts.
These numerous cases of the occurrence of what are otherwise Eastern groups in West Africa, undoubtedly suggest some correspondence of physical conditions which renders this portion of the continent alone suitable to them. The further question, how they got there at all, is elucidated by what we know of the past history of Africa and Europe. It is now generally admitted that, before the Miocene period, Tropical and South Africa was cut off from the great continent of the northern hemisphere by a wide arm of the sea. It was then in fact an island, or perhaps a group of large islands, and probably
contained only some of the lower forms of mammalian life, among which the lemurs and the insectivora were conspicuous. When, during the Miocene period, it became united to Europe and Asia, it was at once overrun by a number of the large mammalia of that continent, such as elephants, rhinoceroses, lions, giraffes, antelopes, hippopotami, apes, and many other forms whose remains are found in abundance in the Miocene deposits of France, Germany, Hungary, Italy, and Greece. South Europe must have then possessed a sub-tropical, if not a tropical, climate, the gradual deterioration of which led to the extinction of most of these animals, while in Africa they have survived and greatly multiplied. Among the European fossils of this period we find some of especial interest. Such are, the genus Hyomoschus, mentioned by Dr. Sclater as one of the Indian group of chevrotains still living isolated in West Africa; a monkey, closely allied to Colobus and Semnopithecus and perhaps an ancestral form of these two groups which are now separated in Asia and West Africa. Among birds, we have remains of the snake-eating secretary bird; Necrornis, supposed to be allied to the peculiarly African plantain-eaters; and a parrot allied to the well-known grey parrots of Africa. Here, then, we have traced the origin of the resemblances between certain animal forms of tropical Asia and West Africa to their having been derived from a common source in the great northern continent, and we are justified in supposing that the Pitta angolensis is also a descendant of one of these Miocene forms. A species of Pitta even now inhabits Japan, so that we may well suppose the genus to have originated in Europe or Western Asia in the warm Miocene period. It was probably once more abundant in Africa, but, along with the Hyomoschus and the crested hornbill, is now in process of extinction in the one continent, while its allies continue to flourish in the other.
The Problem of Allied Forms in all the Tropical Regions.
Leaving for the present the next three cases of difficulty adduced by Dr. Sclater, we will pass on to the
T 2
subject of allied forms occurring in the tropics of both hemispheres. Besides the barbets (Megalæmidæ) which occur in the tropics of Asia, Africa, and America in almost equal abundance, we have the trogons, abundant in Asia and America, but with a single peculiar genus of two species in Africa, and the tapirs confined to the Malay islands and tropical America; while equally remarkable are two genera of snakes, Dryiophis and Dipsadoboa, confined to West Africa and tropical America. Towards an explanation of these curious anomalies we have the very interesting fact, that tapirs closely resembling those now living abounded in Europe during the Miocene period, and continued to live in the Pliocene period, both in France and England, as well as in North America. This suggests that a tropical climate is not essential to these animals, and that their present restricted range is due to other than climatal causes. We may also be sure that if they could live so far north as our island in the Pliocene period, they might have ranged considerably further north during the earlier and warmer Miocene. The only difficulty is, how did these Miocene tapirs reach America? and if we can find any reasonable answer to this question we may consider that it will equally apply to all the other cases which have been mentioned.
The close relation between many of the extinct mammalia of North America and Europe at successive periods, while in other cases entire groups have always been restricted to one continent only, renders it certain that there existed at several distinct epochs some land connection sufficient to enable terrestrial animals to pass between them. The sea at Behring Strait is so shallow that we may safely conclude that the continents of Asia and America have here been recently connected, while the shallow Okhotsk, Japan, and Yellow seas indicate a large extension of the lowlands of Eastern Asia; but the very deep ocean comes up to beyond 55° N. latitude on the east side of Kamschatka, so that this part of the connecting land would probably always have had a warm temperate rather than a tropical climate. On the European side we find between the west of Ireland and Newfoundland a
maximum depth of about 12,000 feet, but with large areas between 5,000 and 10,000 feet deep; and although this implies an immense subsidence, it is not very improbable that all the area from this line northward to Greenland and Iceland was dry land during some part of the Miocene period. In support of this view it may be noted that the Alps, the Pyrenees, the Rocky Mountains, and even the Himalayas, were all in early Miocene times many thousand feet lower than they are now. This is proved by the fact of Eocene and Miocene marine deposits of great thickness, which must have been formed in rather deep water, being found elevated from ten to sixteen thousand feet above the sea-level. As an example we may mention the Dent du Midi in Switzerland, where marine shells of early Miocene or late Eocene type are found at an elevation of 10,940 feet; and, as this mountain must have suffered enormous denudation, these figures can only represent a portion of the rise of the land, most of which occurred during the Miocene period. To balance this rise over extensive areas on both sides of the Atlantic, there must have been corresponding areas of subsidence, and we may fairly locate these where the indications of palæontology and geography concur in rendering them probable. We have already seen that the migrations of mammalia between Europe and America have been such as to render some land route necessary,1 while the broken-up character of the coasts of Ireland and Newfoundland, Labrador, Greenland, and Iceland, with the extensive bank of the Azores, all point to a certain amount of recent sinking of land on the outskirts of this area of great depression.
To Dr. Sclater's question—Where did the tropical land exist which afforded the passage of the tropicopolitan forms from one continent to the other?—it may therefore be answered: It existed in the north temperate zone during some part of the Miocene period, at the time probably when a rich warm temperate flora covered what are now the icy wastes of Greenland and Spitzbergen. In the North Atlantic a continuous land may have united
1 For particulars of these migrations see the writer's Geographical Distribution of Animals, vol. i. pp. 140, 153.
Europe and America at about the latitude of London, without implying a greater amount of subsidence than would balance the elevation which we know has occurred over extensive areas in Europe and America. We also know that two of the most characteristic tropicopolitan forms—the tapirs and the trogons—existed in Europe in Miocene times; and every geologist will admit that there must have been many others, especially among birds and reptiles, whose remains we have not yet discovered, and never may discover. The transmission of similar forms to tropical Africa and Asia has already been explained in reference to Pitta angolensis; and thus, it appears to me, the problem of tropicopolitan forms is completely solved without making any assumptions but such as are warranted by admitted geological and palæontological facts. It has been necessary to treat the question broadly and to omit many details which require fuller elucidation. I can only now call attention to the obvious fact that the geological age of the remains of any animal type in a given area cannot be held to denote the period of its earliest appearance in that area by migration or otherwise, because, till it became somewhat abundant, there would be little chance of its remains being preserved or discovered. This will apply to the case of the tapirs, which are supposed to have migrated to North America in the Miocene period, but whose fossil remains are not found in any deposits earlier than the Pliocene.
Relations between the West Indies and Madagascar.
We will now return to Dr. Sclater's fourth problem, that of the occurrence of the curious insectivorous mammal, Solenodon, in the Antilles, while its nearest allies are to be found in Madagascar. By the help of the conclusions we have already arrived at, much of the marvel and difficulty of this curious case of geographical distribution vanishes. It is simply an extreme instance of a family of animals which has been long dying out, but which maintains a lingering existence in two remote island groups where it is comparatively free from the competition of higher types,
and where the general physical conditions are favourable. The fact that the family Centetidæ consists of six very distinct genera (five in Madagascar and one in the Antilles) is a sufficient indication that it was once an extensive group. In the Lower Miocene of Auvergne, the fossil remains of a small animal have been found, which is provisionally classed in this very family; and both in Europe and America a considerable number of the remains of Insectivora of peculiar genera have been discovered, indicating that this order of mammals is a very ancient one, which probably long ago arrived at its maximum of development, and has been diminishing in proportion as the larger and more perfectly organised forms have been increasing. It is interesting to note that the two localities where the Centetidæ still linger have many remarkable similarities and correspondences. Both are insular groups of the first rank; both are separated from their adjacent continents by very deep sea; both are situated just within the line of the tropic; both are subject to hurricanes; both are very mountainous; in both all the higher mammalia are very deficient; and the differences of their forms of life from those of the adjacent lands are such as to indicate that they have both remained insulated for a considerable period geologically. There can be little doubt that these resemblances have something to do with the continued existence in both of isolated members of a once widespread group of mammals, of a comparatively low type of organisation, and unable to bear the competition to which they have been exposed in continental areas. The same principles will, of course, explain the presence in Madagascar of a mouse allied to an American group, of three American genera of colubrine snakes, and of lizards belonging to the peculiar American family Iguanidæ, as well as of the beautiful green diurnal moths of the genus Urania, and several beetles of decidedly South American affinities. In some other cases we have, as it were, a relic of the former wide extension of now restricted groups. Thus, one genus of snakes, Ahætulla, is found in Africa and South America as well as in Madagascar; while a genus of geckoes, Phyllodactylus, inhabits
also America and Australia, and there are many similar cases among insects.
After what has been now advanced, the distribution of the lemurs (which forms Dr. Sclater's fifth case) will offer little difficulty. Every indication points to this being a group of great antiquity, and to its having been once very widely spread. Its still existing remnants are scattered from Sierra Leone to Celebes, and from Natal to Eastern Bengal and South China; and they are so varied that they require to be classed in three distinct families and thirteen genera. Still more important is the proof of their extreme antiquity afforded by the recent discovery, in the Eocene deposits of the South of France, of a skull of an unmistakable lemur, allied to one of the still living forms of West Africa known as the "Potto"; while several other fossils of the same age are also believed to belong to the lemurine group. In North America, too, abundant remains have been found in the Lower Eocene deposits, which are believed to be intermediate between lemurs and the South American marmosets. This clear evidence both as to the antiquity and the wide range of the lemurs renders it quite unnecessary to postulate any special changes of sea and land to account for their actual distribution. Inhabiting Europe in Eocene times, they were probably spread over the whole northern continent, and would as easily migrate southward into their present habitats as the hedgehogs, the civets, the chevrotains, or the porcupines, which have all a somewhat similar, but far more extensive distribution. Like the Centetidæ, the lemurs find Madagascar best suited to them, more no doubt from the absence of competitive forms than from any peculiar physical conditions. On the great continents they are usually scarce, and are protected by their nocturnal habits and by frequenting dense forests. They thus continue to survive in the midst of creatures of a higher type and more recent origin than themselves, and, together with the opossums of America and some of the smaller marsupials of Australia, seem to have handed down to us a sample of the forms of life which flourished in the earliest tertiary or even in mesozoic times.
Distribution of Gigantic Land-tortoises.
The last case of anomalous distribution—that of the giant land-tortoises of the Mascarene and Galapagos islands—offers perhaps less real difficulty than any of the preceding, on account of the existing wide range and the extreme antiquity of the genus Testudo to which they all belong. This genus occurs in the Miocene deposits of Europe and India, and in the Eocene of North America; and living species are abundant in Africa and Asia, and are also found in South Europe and North America. It has evidently, therefore, been a dominant group during a large portion of the tertiary epoch, and it still maintains a vigorous existence. There does not seem to be any evidence that these giant species of the two hemispheres are more closely allied to each other than the smaller forms of remote regions; for though the Galapagos tortoises and the extinct species of the Mascarene islands both belong to a flat-headed type, they may have differed in important external characters. Their gigantic size is evidently due to their seclusion for countless generations in islands where they were entirely free from the attacks of enemies, and where they could procure abundance of food; both natural and sexual selection giving the advantage to the larger and stronger individuals. The only difficulty is how they reached the Galapagos. But as we may go back to the middle of the tertiary epoch for this event, it is not an improbable supposition that the coast of South America then extended considerably westward, while the islands themselves may have been more extensive, thus reducing the dividing strait to a width across which either the adult animals or their eggs might be floated by currents or surface-drifts. Their entrance to the Mascarene islands from Africa might have been effected in a similar manner. This is the solution suggested by Dr. Günther himself,1 and it is one which perfectly harmonises all the known facts.
1 Nature, vol. xii. p. 297.
Closely Allied Forms in the Same Area.
The only other difficulty suggested by Dr. Sclater rather applies to the theory of natural selection itself than to geographical distribution. He asks how we are to account for closely allied forms so frequently inhabiting the same area, while in so many other cases allied species are strictly limited to distinct areas, to the diverging physical and organic conditions of which they are supposed to be adapted. We think that Dr. Sclater has himself furnished a clue to one mode of solution, in his statement that the willow-warbler and the chiff-chaff, though so alike externally, are yet quite distinct in mode of life and habits. It is evident, that a variation in the habits of a portion of the individuals of a species would lead to their mutual association and at the same time to their separation from the parent form, and would thus obviate that tendency to the intercrossing of the different varieties which would undoubtedly occur if the variation were one of colour or form only. Is it not probable, therefore, that where two or more closely allied species inhabit the same area, they have arisen at first by means of useful variations of instincts or habits; while those which inhabit adjacent but separate areas may have arisen by favourable variations of colour, form, or constitution only? In support of this view it may be noted that the coal and marsh titmice, which are very closely allied, differ considerably in habits; while the great and blue titmice, which are very different in external characters, agree closely in habits, and are often seen together.
Summary of General Principles.
Returning to the general question of zoological distribution and its anomalies, it has been shown, I trust, that the only mode of explaining the existing distribution of living things is by a constant reference to those comparatively slight but often important changes of sea and land, which the most recent researches show to be alone probable; and, what is still more important, by recognising
the undoubted fact that every group of animals whose distribution is discontinuous is now more or less in a fragmentary condition, and has, in all probability, once had a much more extensive range, to which its present distribution may offer no clue whatever. Who would ever have imagined, for example, that the horse tribe, now confined to Africa and Asia, formerly ranged over the entire American continent, north and south, in great abundance and variety; or that the camel tribe, now confined to Central Asia and the Andean region of South America, formerly abounded in North America, whence in fact our existing camels were almost certainly derived? How easy it is to imagine that analogous causes to those which have so recently exterminated the horses of America and Europe might have acted in a somewhat different direction, and have led to the survival of horses in South America and Africa, and their extermination elsewhere. Had this been the case, how strong would have been the argument for a former union of these two continents; yet we now know that these widely separated species would be merely the relics of a once dominant group which had occupied and become extinct in all the northern continents.
Discoveries of extinct forms remote from the countries they now inhabit, are continually furnishing us with new proofs that the great northern continents of the two hemispheres were really the birthplace of almost if not quite all the chief forms of animal life upon the globe; while change of climate, culminating in the glacial epoch, seems to have been the motive power which has driven many of these forms into the tropical lands where they now alone exist.
If we give full weight to these various considerations, and at the same time bear constantly in mind the extreme imperfection of our knowledge of extinct land animals, we shall, I believe, have no difficulty in explaining most of the apparent anomalies in zoological distribution, and in imagining a possible and even probable solution for those extreme cases of difficulty which the facts at our command do not yet permit us to explain in detail.
Let us now briefly summarise the general principles on which the solution of problems in zoological distribution depends.
During the evolution of existing forms of animal life, we may picture to ourselves the production of successive types, each in turn increasing in variety of species and genera, spreading over more or less extensive regions of the earth's surface, and then after arriving at a maximum of development, passing through various stages of decay, dwindling to a single genus or a single species, and finally becoming extinct. While the forms of life are thus, each in turn, moving on from birth to maturity and from maturity to decay and death, the earth's surface will be undergoing important physical changes, which will sometimes unite and sometimes separate contiguous continents or islands leading now to the intermingling, now to the isolation, of the progressing or diminishing groups of animals. Again, we know that climates have often changed over a considerable portion of the earth, so that what was at one time an almost tropical region has become at another time temperate, and then even arctic; and these changes have, it is believed, been several times repeated, leading each time to important changes, migrations, and extinctions of animal and vegetable life.
It is by the combined effect of these three distinct sets of causes, acting and reacting on each other in various complex ways, that those curious examples of erratic distribution of species and genera have been produced which have been so long a puzzle to the naturalist, but which have now, it is believed, been shown to be the natural and inevitable results of the process of animal development, combined with constant changes in the geography and in the climate of the earth.
CHAPTER XIV
THE ORIGIN OF SPECIES AND GENERA1
THE meaning of the term—now become a household word in science—"the origin of species," is often entirely misapprehended. It is very generally thought to mean the origin of life and of living things, and people are surprised and almost incredulous when told that Mr. Darwin himself, in the latest edition of his celebrated work, still refers that origin to divine agency. Such however is undoubtedly the case, as shown by the following passage which concludes the volume: "There is grandeur in this view of life, with its several powers, having been originally breathed by the Creator into a few forms or into one; and that, while this planet has gone cycling on according to the fixed law of gravity, from so simple a beginning, endless forms most beautiful and most wonderful have been, and are being, evolved."1
The mistake above alluded to has arisen from ignorance of the meaning of the word "species," the "origin" of which Mr. Darwin undertakes to show. A species may be defined as a group of individuals of animals or plants which breed together freely and reproduce their like; whence it follows that all the individuals of a species, now living or which have lived, have descended from a few common ancestors, or perhaps from a single pair. Thus all
1 In a letter to Sir Joseph Hooker in 1863, Darwin explains that in using the term "Creator" he really meant "appeared by some wholly unknown process,"—adding—"It is mere rubbish thinking at present of the origin of life; one might as well think of the origin of matter" (Life and Letters, iii., p. 18.)
horses, whether Shetland ponies, racers, or cart-horses, form one species, because they freely breed together, and are known to have all descended from a common stock.
By the same test the common ass, the kiang, the quagga, and the zebra, are each shown to be distinct species; for though sometimes two of these species will breed together, they do not do so freely, they do not reproduce their like but an intermediate form called a mule, and these mules are not capable of reproducing their kind, as are the offspring of any pairs of a single species. What Mr. Darwin did was to prove, by an overwhelming array of evidence and a connected chain of irresistible argument, that, just as all horses and all asses have each descended from a few common ancestors, so have all asses, horses, quaggas, and zebras descended from a much more remote common ancestral form; and that the same thing has occurred with every group of allied species. This is the "origin of species" by descent with modification, or, in other words, by evolution; while "natural selection" was the term applied to the set of natural causes which formed the motive power and guiding principle by which the change from one species to another was brought about.
In a very few years after the publication of this theory, it had literally extinguished, among all thinking men, the doctrine of special creation which had before largely prevailed; and some, who were its most violent opponents at the outset, now accept the fact of evolution as applied to almost every group of organised beings. At the present day there is perhaps no single naturalist of reputation who upholds that doctrine of the independent origin of each species of animal and plant, which was a very few years ago either tacitly accepted or openly maintained by the great majority of naturalists. Surely no such revolution in scientific thought was ever effected by one man in so short a period!
At first the opponents of Darwinism opposed evolution as well; but of late years the opposition is directed wholly to the adequacy of the causes which Mr. Darwin maintains are sufficient to explain the origin of each species
from some pre-existing species, and therefore the origin of all existing species from some one or more ancestral forms. It is maintained that there are other laws at work besides natural selection, and Mr. Darwin has himself admitted that there probably are such. Most of the opponents of Darwinism argue in favour of some guiding or organising power, either internal or external, as absolutely necessary to the production of the kind and amount of variation necessary for the development of the various complex organs and special adaptations which characterise each important class of animals. Others go still further and maintain that "natural selection" is powerless to produce new species in any case, its function being to keep those which are produced in a state of health and perfection by weeding out all that are imperfect; or, they argue that, so long as the "cause of variation" is unknown the power that preserves those variations when they have arisen plays a very subordinate part. These last writers maintain that the causes, whatever they are, which produce certain variations in certain species at certain times, are the true and only causes of the origin of species.
Now all these objections, in so far as they refer to the origin of the different species of one genus from a common ancestral species, or even of all the species and genera of one family from some still more remote ancestor, may, I think, be shown to be invalid; because we have direct evidence, almost amounting to demonstration, that changes to this extent are producible by the known laws of variation and the admitted action of natural selection. But when we go further back, and propose to account for the origin of distinct families, orders, and classes of animals by the same process, the evidence becomes far less clear and decisive. We find groups possessing organs no rudiment of which exists in other groups; we find classes differing radically in structure from other classes; and we have no direct evidence that changes of this nature are now in progress, as we have that the lesser changes resulting in new species and new genera are in progress.
Yet the evidence that these deeper and more important changes in the structure of organised beings have taken
place by gradual steps through the ordinary processes of generation is overwhelming. The numerous intermediate links that have been discovered both among living and extinct animals, and especially the wonderful community perceptible in the embryological development of the most diverse living types, force upon us the conclusion that the entire animal and vegetable kingdoms owe the wonderfully diversified forms they now exhibit to one unbroken process of "descent with modification" from a few primeval types. It is indeed generally assumed that if we go so far, we must admit one original type of living organism; but this does not seem necessary. By means of whatever laws we suppose living things first to have originated, why should not the primeval germs have appeared several times independently, and in forms determined or modified by the infinitely varied chemical and physical conditions to be found in the crust of the earth? The identity of ultimate structure and the wonderful similarities of development of all organisms may be due to the unity of the laws by which organic life was first produced; the diversity of the great types of animal and vegetable forms may be due to the operation of those laws at different places, acting on different combinations of elements, which are subject to unlike physical conditions.
The point here insisted upon is, that the origin of all organisms, living and extinct, by "descent with modification," from one primeval germ or cell, is not necessarily the same thing, and is not included in, "the origin of species by means of natural selection." The latter we not only know has occurred, but we can follow the process step by step by means of known facts and known laws; the former, we are almost equally certain, has occurred, but we cannot trace its steps, and there may have been facts and laws involved of which we have no certain knowledge. The terms "laws of growth," "laws of development," "laws of inheritance," "laws of variation," "laws of correlation," "direct action of the environment," "laws of habit and instinct," with some others, are used to express the action of causes of which we are almost wholly ignorant, as we are of the nature of life itself. Now Mr.
Darwin has himself admitted that there are these unknown causes at work, and that "natural selection is the most important but not the exclusive means of modification." There may be some question as to the term "most important," if, as is not improbable, the most radical differences in animals and their most important organs could not have been produced by it alone in the same way as the specific modifications of a genus or family may be produced. This, however, is a fair matter for discussion and research, and will probably continue to be so for many generations; and even if it should be ever proved that higher laws than "natural selection" have brought about the more fundamental divergences of the animal and vegetable kingdoms, this will not be held to detract in any way from the greatness and the value of Mr. Darwin's work, any more than it will be held to detract from the greatness of Newton, if it should some day be demonstrated that the law of gravitation as expressed by him is not absolutely true, but that (as some physicists now suppose) it should be found to be subject to a higher law for remote stellar distances, or in the case of such exceptional phenomena as those presented by comets.
No thoughtful person can contemplate without amazement the phenomena presented by the development of animals. We see the most diverse forms—a mollusc, a frog, and a mammal—arising from apparently identical primitive cells, and progressing for a time by very similar initial changes, but thereafter each pursuing its highly complex and often circuitous course of development, with unerring certainty, by means of laws and forces of which we are almost totally ignorant. It is surely a not improbable supposition that the unknown power which determines and regulates this marvellous process may also determine the initiation of those more important changes of structure and those developments of new parts and organs which characterise the successive stages of the evolution of animal forms, In so far as Mr. Darwin denies the necessity of any such power, and maintains that the origin of all the diverse forms and types and all the complex structures of the organic world are due to identi-
VOL. I. U
cally the same laws and processes as are adequate to produce the different species of Rubus or of Canis, from some ancestral bramble or dog respectively, his opponents have undoubtedly a case well worthy of being argued out in the courts of science. They should, however, remember that no final judgment has been given or can be given while the evidence on both sides is not only circumstantial but imperfect and contradictory; and it would be well not to declare too confidently that Mr. Darwin's theory has hopelessly broken down, since a majority both of naturalists and geologists, whose evidence as experts will undoubtedly have great weight with the educated public, are at present altogether in his favour.
How New Species have Originated.
Leaving this great case to be discussed and argued in weighty volumes by specialists in science, I here propose to deal briefly with that much smaller but still important question, of the origin of the species of a genus or of a family—that is, of groups of organisms differing, as the wolf, dog, and fox differ, among animals, or as the numerous species of oaks or of primulas differ, among plants; and I hope to be able to show that in these cases there is hardly any room for doubt as to the mode in which the change from species to species has been effected.
We have to inquire, then, how it is that new species have arisen, supposing the world to have been then very much as it is now; and what becomes of them after they have arisen. In the first place we must remember that new species can only be formed when and where there is room for them. If a continent is well stocked with animals and plants, there is a balance between the different species, those best adapted to the varied existing conditions maintaining themselves in the largest numbers, while others, being only adapted to special conditions that occur in limited areas, are far less numerous; the former are common and widespread, the latter rare or local species. If the entire group of organisms in any country has existed for a sufficient time to have been subjected to all the
varying conditions which occur during considerable cycles of climatal and other changes, the balance will have become well established, and so long as no change takes place in the conditions no new species will arise.
But now let us suppose some change to begin, either of climate or geography. The land may sink or it may be elevated, in the former case diminishing in area and perhaps becoming divided by an arm of the sea, in the latter case increasing in area and perhaps becoming united with extensive lands formerly separated from it; or the climate may become moister or drier, hotter or colder, more extreme or more equable, and any one of these changes or any combination of them would, it is easy to see, produce a special effect on the forms of life. The vegetation would in almost any case become changed, and this would affect both the insects and the higher animals in a variety of ways. New enemies or new competitors might be admitted, and these would certainly cause the extermination of some of the rarer species, and perhaps greatly reduce the numbers of those which had been most numerous. Others might, from the same general causes, obtain fresh supplies of food, or have fresh areas over which to spread themselves opened to them.
These are the first and most obvious effects of such changes, but there are others still more important, and not less certain to be produced. We have supposed each of the species which inhabited the country to be well adapted to the conditions of its existence, to be able to obtain food for itself and young, to protect itself against all kinds of enemies, and to be able to resist the ordinary inclemencies of the seasons, and to do all this in competition with the numerous other species by which it was surrounded. But now, we suppose, all these conditions and surroundings to be undergoing change, either at the same time or in succession; and, in order to become equally well adapted to the new conditions, some of the species will require to undergo a corresponding change, either in structure, habits, colour, or some other characters. New enemies may necessitate greater swiftness, or greater cunning, or less conspicuous colours; less
U 2
abundant food may necessitate some modification in structure better adapted to secure it, or the means of ranging over a wider area to search for it; while a severer climate may necessitate a thicker covering, or more nourishing food, or new kinds of shelter. To bring about these changes, "variation" and the "struggle for existence" come into play. Each year the forms less adapted to the altered conditions die out, while those variations which are more in harmony with them constantly survive; and this process, continued for many hundreds or thousands of successive generations, at length results in the production of one or more new species.
Variability.
We now come to the difficulty which has been repeatedly put forward, and which seems very great to all who have not studied groups of species as they occur in nature, and which is expressed in the question "How comes it that variations of the right kind and sufficient in amount have always occurred just when they were wanted, so as to form the endless series of new species that have arisen?" and it is more especially to answer this question that the present chapter has been written.
Few persons consider how largely and universally all animals are varying; yet it is certain that if we could examine all the individuals of any common species we should find considerable differences, not only in size and colour but in the form and proportions of all the parts and organs of the body. In our domesticated animals we know this to be the case, and it is by means of the continued selection of such slight varieties to breed from that all our extremely varied domestic animals and cultivated plants have been produced. Think of the difference in every limb, in every bone and muscle, and probably in every part, internal and external, of the whole body, between a greyhound and a bull-dog! Yet, if we had the whole series of ancestors of these two breeds before us, we should find them gradually converge till they reached the same original type, while between no two successive generations would there be any greater difference
than now sometimes occurs in the same litter. It is often thought, however, that wild animals do not vary sufficiently to enable any such change as this to be brought about in the same limited time; and though naturalists are well aware that there is little, if any, difference in this respect between wild and domesticated species, it is only recently that they have been able to adduce positive proof that this is the case.
We owe this proof to an American naturalist, Mr. J. A. Allen,1 who was the first to make a series of observations and measurements of the mammals, and more especially of the birds, of the United States; and he found a wonderful and altogether unsuspected amount of variation between individuals of the same species even when inhabiting the same locality. They differ in the general tint, and in the distribution of the colours and markings; in general size, and in proportions; in the length of the head, feet, wings, and tail; in the length of particular feathers, thus altering the shape of the wing or tail; in the length of the tarsi and of the separate toes; and in the length, width, thickness, and curvature of the bill. These variations are by no means small in amount or requiring very accurate measurements for their detection, since they often reach one-seventh, one-sixth, or sometimes even one-fourth, of the entire average dimensions. Thus, in twelve species of small birds, the variation in twenty-five or thirty specimens, all taken in the same locality, and of the same sex and age was, in the length of the folded wing, from 14·5 to 21 per cent., and in the length of the tail from 14 to 23·4 per cent. If we take individual cases, we find equally striking facts. Wilson's thrush (Turdus fuscescens) was found to vary in length of wing from 3·58 to 4·15 inches, and in the tail from 3·55 to 4 inches. In the Blue-bird (Sialia sialis) the middle toe varied from 0·77 to 0·91 inch, and the hind toe from 0·58 to 0·72 inch; while the bill varied from 0·45 to
1 "On the Mammals and Winter Birds of East Florida; with an Examination of certain assumed Specific Characters in Birds, and a Sketch of the Bird Faunæ of Eastern North America." By J. A. Allen. (Bulletin of the Museum of Comparative Zoology at Harvard College, Cambridge, Mass., vol. ii. No. 3.)
0·56 inch in length, and from 0·30 to 0·38 inch in width. In the Yellow-crowned Warbler (Dendrœca coronata) the quills vary in proportionate length, so that the first, the second, the third, or the fourth, is sometimes the longest; and a similar variation of the wing, involving a change of proportion between two or more of the feathers, is recorded in eleven species of birds. Colour and marking vary to a still greater extent. The dark streaks on the under parts of the American Song-sparrow (Melospiza melodia) are sometimes reduced to narrow lines, while in other specimens they are so enlarged as to cover the greater part of the breast and sides of the body, sometimes uniting on the middle of the breast into a nearly continuous patch. In the small spotted Wood-thrushes (of the sub-genus Hylocichla) not only does the general tint of different parts vary greatly, but this is accompanied by great variation in the markings, some specimens being very pale with in distinct narrow lines on the breast, while others have dark plumage and dark, broad, triangular markings. It must be remembered that all these differences are independent of those due to age, sex, season, or locality, and consist solely of what may be termed the normal individual variation of the species.
It is, however, often supposed that variations occur at any one time in single characters only, all the rest remaining invariable; and it is objected that to adapt a creature to new conditions it must be modified in several ways at once. But a reference to the tables given by Mr. Allen shows that this coincident variation of several characters does exist to a remarkable extent. He has given the variation of no less than nineteen characters in ten species of birds, from a comparison in each case of only twenty specimens, all of the same sex, all fully adult, and all taken in the same localities. On marking the specimens which have each character at a maximum or minimum development, we find the most curious combinations. We find, for example, that the largest specimens have not always the longest wings or tails, or the smallest specimens the shortest; the proportion of the different parts of the wing varies quite regardless of the actual dimensions; the
length of any toe varies independently of the length of the tarsus; a long head sometimes goes with a short, sometimes with a long wing; while the width of the bill seems to vary independently of its length or of any of the other parts of the body. All these variations, too, are very considerable in amount. Thus among twenty male Baltimore Orioles the total length varied from 7 to 8 inches; the wing from 3·45 to 3·85 inches; the tail from 2·70 to 3·10 inches; the primaries extended beyond the secondaries from 0·56 to 0·90 inch; the tail extended beyond the upper coverts from 1·37 to 1·87 inches; the tarsus varied from 0·83 to 1·02 inch; the hind toe varied from 0·62 to 0·75 inch, and the middle toe from 0·82 to 1·00 inch; the head varied in length from 1·50 to 1·62 inches; the beak in length from 0·74 to 0·84 inch, and in width from 0·32 to 0·38 inch. And if these differences and these combinations, indicating many diverging proportions between two or more characters, are found among only twenty specimens, we may certainly expect much greater differences in every character, and these differences combined in an endless variety of ways, among the millions of individuals which constitute every common species. Not only, therefore, is it clear that there is, among birds at all events, ample individual variation for natural selection to work upon, but, what is even more important, that coincident variations in every conceivable combination are also available.
Among mammalia we have fewer materials for comparison, but there is good reason to believe that they are quite as variable as birds, if not even more so. Among twenty males of the Grey Squirrel, whose dimensions are given by Mr. Allen, we find the length of the tail to vary as 3 to 4, of the fore foot as 9 to 11, and of the hind foot as 6 to 7. The Virginian Opossum also varies greatly in colour, and in the size and proportions of all the parts, including the skull, the variation amounting to nearly twenty per cent.1
1 A number of diagrams illustrating these and many other facts of variation, are given in my Darwinism (Chap. III.), where the subject of the nature of variation is more fully discussed.
If now we consider the population of a species with regard to any particular character or combination of characters, we may divide it into three groups—a central group in which the mean or average development prevails with little variation, one in which the character is excessively, and one in which it is little developed. These groups would not be of equal extent, the central portion—that in which the mean characteristics prevailed—being, in accordance with the law of averages, much more numerous than the extremes; perhaps twice or even three times as great as either of them, and forming such a series as the following:—Maximum development 10, mean 30, minimum 10. These figures, whatever their exact proportions, would probably be pretty constant, for we have no reason to believe that the mean characters, or the amount of variation of a species, change materially from year to year or from century to century; and we may therefore look upon the central and most numerous group as presenting the typical form of the species, being that which is best adapted to the conditions in which it has actually to exist, while the extremes, being less perfectly adapted, are continually weeded out by natural selection.
Besides the individual variation above noticed in birds of the same locality, another set of variations appears in birds of the same species inhabiting different localities. In North America birds decrease in size as they inhabit localities further south, while they become larger as we go north. In mammalia, on the other hand, there is generally a decrease of size both north and south from a central position where the species is at a maximum. Strange to say, the bill of most birds increases in length towards the south, sometimes relatively, but in other cases absolutely, so that the smaller southern birds sometimes have a bill actually longer than the northern larger individuals. This peculiarity occurs in the genera Quiscalus, Agelæus, Troglodytes, Seiurus, &c., and is illustrated by numerous figures in Mr. Allen's work. In some cases, as in the American crow, the bill is so much larger in the south that the Florida birds have been recognised as a distinct named variety.
Colour also varies greatly in correspondence to latitude and longitude. Dark-coloured birds are said to become blacker towards the south; in others the yellow or red bands become deeper; while in those transversely banded, the dark bands become broader, and the light ones narrower. Those with white spots or bands have them smaller in the south, and sometimes lose them altogether. These differences are sometimes so great that the extreme northern and southern forms might be considered distinct species were it not for the perfect gradation of intermediate types in the intervening localities. There is also an increase of intensity of colour from east to west, as exhibited by the same or by closely allied representative species inhabiting the Atlantic and Pacific coasts respectively. In the desert plains of the interior, however, the colours are paler than on either coast; but this is no doubt a protective modification, assimilating the tints of animals to the rock or surface soil on which they dwell. In some cases well-marked varieties of the same species appear to be confined to the Eastern States and to California respectively, as, for example, the eastern and western forms of Bewick's Wren (Thryothorus Bewickii), which differ greatly in the length of the bill, although otherwise almost identical; and as these two forms do not, so far as yet known, anywhere intermingle, they afford a good example of the first step in the formation of a new species. The beautiful purple finch (Carpodacus purpureus) of the Eastern States, and its western form, which has been named californicus, perhaps form another example; but until the range of these birds is fully and accurately determined, we cannot be sure that there is not some limited area where the two forms intermingle and their distinctive characters disappear.
From the fact of variation, so extensive as regards the number of variable characters and so large in absolute amount as has now been proved to exist in many species, we may fairly draw the conclusion that analogous variations, sometimes of less and sometimes of greater extent, is a general characteristic of animals in a state of nature; and with such materials to work with, it becomes easy
to understand how new species may arise. For example, the peculiar physical or organic conditions that render one part of the area occupied by a species better adapted to an extreme variety may become intensified. The most extreme variations in this direction will then have the advantage, and will multiply at the expense of the rest. If this change of conditions should extend over the whole area occupied by the species, this one extreme form will replace all the others; while, if the area should be cut in two by subsidence or elevation, the conditions of the two portions may be modified in opposite directions, each becoming adapted to one extreme form. The original type of the species will then have become extinct, being replaced by two species, each distinguished by a combination of certain extreme characters which had before existed in some of its varieties.
Effect of Changed Environment.
The changes of conditions which lead to such selection of varieties are very diverse in their nature; and new species may thus be formed diverging in many ways from the parent stock. The climate may change from moist to dry, or the reverse, or the temperature may increase or diminish during long periods, in either case requiring some corresponding change of constitution, of covering, of vegetable or of insect food—to be met by the selection of variations of colour or of swiftness, of length of bill, or of strength of claws. Again, competitors or enemies may arrive from adjacent countries, giving the advantage to such varieties as can change their food, or by swifter flight or greater wariness can escape their new foes. In this way several series of changes may occur, each brought about by the pressure of changed conditions; and thus what was before a single species may become transformed into a group of allied species, differing from each other in a number of slight characters, just as we find them in nature.
How Genera Originate.
Let us now see how the same principles will explain the origin of genera. A genus is a group of allied species which differs from all other groups in some well-marked characters of a structural rather than of a superficial nature. For example, species of the same genus usually differ from each other in size, in colour or markings, in the proportion of the limbs or other organs, and in the form and size of such superficial appendages as horns, crests, manes, and the accessory ornamental plumes of birds; but they generally agree in the form and structure of important organs, as the teeth, the bill, the feet, and the wings. But when two groups of species differ from each other constantly, and to a well-marked degree, in one or more of these latter characters, they are said to belong to distinct genera; and we have seen that species vary in these as well as the more superficial characters—the bill, the feet, and the wings varying in size and proportions just as frequently as do the colours or the ornamental parts of the plumage. If then, in any portion of the area occupied by a species, some important change of habits becomes useful to it, all such structural variations as facilitate the change will be accumulated by natural selection, and when they have thus acquired the proportions most beneficial under the altered conditions, we shall have the first species of a new genus.
A creature which has been thus modified in important characters will form a new type, specially adapted to fill its place in the economy of nature. It will almost certainly have arisen from an extensive or dominant group, because such only are sufficiently rich in individuals to afford an ample supply of the needful variations, and it will therefore inherit the vigour of constitution and adaptability to a wide range of conditions which gave success to its ancestors. It will thus have every chance in its favour in the struggle for existence; it may spread widely and displace some of its nearest allies, and in doing this will extend into new areas, where it will be
subject to a somewhat altered set of conditions, and by further variation and selection may become the parent of a number of subordinate species. It will now have become a dominant genus, occupying an entire continent, or perhaps even two or more continents, spreading on all sides till it meets with competing forms better adapted to the conditions which there prevail.
Such a genus may continue to exist during long geological epochs. Bats of the genus Vespertilio lived in the Eocene period, and still range over all the globe, while fossil land shells of the genus Pupa, hardly distinguishable from some now living, are found in the ancient carboniferous deposits. Generally, however, a time comes to every genus when either physical changes, or competing forms, or new enemies, are too much for it, and it begins to lose its supremacy. First one, and then another of its species dwindle away and become extinct, till at last two or three only remain. Sometimes these soon follow the others, and the whole genus dies out, as thousands of genera have died out during the long course of the earth's life-history; but it also often happens that a few species continue to maintain themselves in areas where they are removed from the influences that have exterminated their fellows. Thus the mudfish of Queensland (Ceratodus Forsteri) and the Trigonia, a bivalve mollusc of the Australian seas are the only living representatives of genera which lived in the Triassic period.
I have now, I think, shown that one of the most general objections to natural selection as producing new species—namely, that there are enormous chances against the right kind of variations occurring just when they are required—is utterly fallacious, by proving that there is ample variation of every kind constantly occurring among animals in a state of nature. It has also been shown that many different kinds of variation are occurring at the same time, and in endlessly varied combinations, so that any required combination of characters could be selected as well as any single character. And when we consider the extreme slowness of almost all the changes of conditions
which lead to the selection of new forms, and the enormous selecting power brought to bear owing to the rapid increase and corresponding great annual mortality among all animals, it is impossible to doubt that the means are adequate to the result. To bring these means clearly before our readers, let us suppose that a pair of birds produce every year six young, and that they live for five years. We thus have thirty birds out of which to replace the two, so that, on the average, at least twenty-eight must die during this time, and many more if any of these live to breed along with their parents. This gives us, as a minimum, a destruction every five years of fourteen times as many birds as exist at any one time. Now let us suppose a change going on which renders it beneficial for a species to obtain longer wings in order to escape from some enemy, and a stronger bill to enable it to capture some fresh insect, both of which (the enemy and the insect) are gradually increasing in the country. Variations of both these kinds occur in abundance every year, to an amount measured by ten or twenty per cent. of the average dimensions. Either of the variations would be useful and would be preserved separately, while the combined variation would be doubly useful and would also be preserved whenever it appeared. A race in which these two characters were from ten to twenty per cent. above the average would therefore be easily produced in twenty or fifty years; while in a thousand or five thousand years a change amounting to thirty or forty per cent.—far greater than distinguishes many species—would probably be brought about. This illustration, I think, renders it clear that the extreme slowness of the action of natural selection on which Mr. Darwin repeatedly dwells, is by no means an essential characteristic of it, but is only due to the fact that physical and other conditions usually change with extreme slowness. But if, as must often have happened, conditions have changed with comparative rapidity, then the enormous amount of individual variation, which would be taken advantage of every year by the survival of the fittest, might effect changes in a single century quite as great as those which distinguish nearly allied species.
Supposed Necessity of showing the Cause of Variation.
Another objection which is dwelt upon with constant reiteration by Mr. Darwin's critics is, that he has not shown the cause of variation, and that whatever it is that causes variation, that is the real "origin of species." This has always seemed to me one of the most unmeaning and irrational of objections, because every explanation must take as a basis well-known facts to explain obscure phenomena. When the geologist explains how the contour of a country has been formed by rain and ice, it is not said that he has explained nothing unless he goes on to show exactly how rain and snow are formed, or even goes further back to the cause of gravitation which is really what gives them all their power to do any work; and when the physicist explains how thunder and lightning are produced by a reference to the electric spark and its accompanying sound, he is not told that the explanation is valueless till he has discovered the nature and cause of electricity itself.
But we may, I think, go further, and say that variation is an ultimate fact of nature, and needs no other explanation than a reference to general principles which indicate that it cannot fail to exist. Does any one ask for a reason why no two gravel-stones or beach-pebbles, or even grains of sand, are absolutely identical in size, shape, surface, colour, and composition? When we trace back the complex series of causes and forces that have led to the production of these objects, do we not see that their absolute identity would be more remarkable than their diversity? So, when we consider how infinitely more complex have been the forces that have produced each individual animal or plant, and when we know that no two animals can possibly have been subject to identical conditions throughout the entire course of their development, we see that perfect identity in the result would be opposed to everything we know of natural agencies. But variation is merely the absence of identity, and therefore requires no further explanation; neither do the diverse amounts of
variation, for they correspond to the countless diversities of conditions to which animals have been exposed either during their own development or that of their ancestors.
This objection has really its only possible justification in the ignorant belief that variations of any tangible amount are rare events occurring at long intervals; and therefore that when any combination of special variations was needed to bring an animal into harmony with changed conditions, the number of individuals varying would not be sufficiently great to prevent their being completely swamped by the typical unvarying forms. Had such been the case, some agency capable of producing a considerable amount of variation when required would undoubtedly have been needed, and this unknown agency might fairly have been claimed to be one of the most important factors in the "origin of species." But now that it is proved by many independent observations, that a large percentage of the individuals of most species vary, in each successive generation, to an amount far greater than is required for natural selection to act upon, the whole difficulty ceases to exist. Variation is seen to be one of the most constant and universal facts of nature, always producing what may be termed the raw materials of species in overflowing abundance, so that, whenever and wherever alteration of the conditions of existence is going on, there is always ready to hand an ample stock of varying organisms, by means of which an almost exact adjustment to those conditions may be kept up.
The facts and arguments now adduced will, it is hoped, enable intelligent readers who are not naturalists to form a clear conception of what is really meant by "the origin of species by means of natural selection," and will satisfy them that the most common and what seem at first sight to be the most weighty objections to it, owe all their force to the ignoring of some of the best established facts in natural history.
I have also attempted to show that the causes which have produced the separate species of one genus, of one family, or perhaps of one order from a common ancestor,
are not necessarily the same as those which have produced the separate orders, classes, and sub-kingdoms from more remote common ancestors. That all have been alike produced by "descent with modification" from a few primitive types, the whole body of evidence clearly indicates; but while individual variation with natural selection is proved to be adequate for the production of the former, we have no proof and hardly any good evidence that it is adequate to initiate those important divergences of type which characterise the latter.1
1 Since this chapter was written the argument has been strengthened by numerous additional observations of variation by other naturalists, many hundreds of individuals of a species being measured; and in every case variation to a large amount has been found to exist in every part or organ thus tested.
CHAPTER XV
A CRITIC OF NATURAL SELECTION ANSWERED
IN Natural Science of July, 1894, the Rev. George Henslow makes some statements with regard to variation and Natural Selection which call for critical remark, especially as they have been treated with great detail in a popular work by the same writer, and are thus likely to spread erroneous views as to Darwin's discoveries, as well as to the general theory of Evolution through variation and the survival of the fittest. He says that, though cultivated plants vary indefinitely, and therefore require selection to produce definite modifications, this is not the case in Nature—"Variation in Nature is always in strict adaptation to the direct action of the environment; in other words, natural variation is always definite." This statement seems to me so extraordinary, and so opposed to well-known facts, that I can only impute it to the use of the terms "vary" and "variation" in two very distinct senses; first, as meaning those individual variations which occur abundantly both in nature and under cultivation; and, secondly, as meaning those particular variations which alone survive under nature and produce either a "variety" or ultimately, a "species." In this latter sense, of course, "natural variation" is definite; but so, in the same sense, are the variations of cultivated plants. From unstable and indefinite "variations" man and nature alike produce definite "varieties." As one out of the innumerable examples of indefinite variation which might be named, are
VOL. I. X
the fifteen different modes of variation observed by Alph. de Candolle on a single oak tree, while in a great number of common species an equal amount of variability may be observed both in wild and cultivated individuals; and all these variations are indefinite, in the sense that they do not usually occur in one direction only, from the typical form. A few examples of such variations have been given in my Darwinism, pp. 76–80. I cannot, therefore, understand either the meaning or the value of the statement—"natural variation is always definite."
It is not quite clear whether Mr. Henslow admits the agency of Natural Selection at all. He says: "I would ask what facts are producible to prove that Natural Selection acts at all on the maintenance, if not the origin, of any floral and, indeed, other structures?" It is, of course, admitted that direct proof of the action of Natural Selection is at present wanting; but the indirect proofs have been so cogent as to overcome the most violent prejudice and opposition, and to convert a large majority of naturalists to a belief in its agency. It is, therefore, rather late in the day to deny its existence without adducing some adequate and proved substitute.
Mr. Henslow's Theory.
Mr. Henslow's contention is that the reaction of vegetable tissues to the environment, is the direct cause of adaptation; and in the special case of flowers he imputes all the variety of form and endless modifications of structure to "the responsive action of the protoplasm, in consequence of the irritations set up by the weights, pressures, thrusts, tensions, &c., of the insect visitors."1 Now the very first essential to this theory is to prove that modifications produced by such irritations are hereditary. Here, if anywhere, we want facts. Yet in the very interesting volume to which Mr. Henslow refers us, crowded as it is with facts and observations, I can find only two or three slight references to this most vital point. At page 147 of the same volume, he quotes Darwin as saying that
1 The Origin of Floral Structures, p. 340.
the excellence of our milking cows and goats may be attributed partly to selection and "partly to the inherited effect of the increased action, through man's art, of the secreting glands," and he adds, "This fact" is strictly analogous to what takes place in the vegetable kingdom! Here we have a mere opinion of Darwin's, nowhere supported by direct observation or experiment and now seriously challenged by a large body of naturalists, set forth as "a fact." Again, at page 157, the case of the various "ant-plants" of the eastern tropics is referred to, and it is stated that Dr. Beccari explains the curious hollow stem in which the ants dwell as partly due to the irritation of the ants inducing hypertrophy of the vegetable tissue, which "then becomes hereditary"; and Mr. Henslow concludes that there is abundant evidence to prove that many organs of a plant, if subjected to irritation, can become materially altered and develop new processes, and, "secondly, that these altered states, if the irritation be persisted in, may become hereditary." Here again are only opinions without a particle of proof; and I can find nothing more to the point in the whole volume. The case of galls is very briefly referred to at p. 144, and their non-heredity is passed by with the remark that the predisposition to produce them may be greater now than formerly, and that the galls themselves may be larger than they were at first. But surely if the effects of insect irritation are anywhere hereditary it would be here. An oak tree which lives several hundred years is subject to this irritation in greater or less degree almost every year, and the irritation itself is not momentary and intermittent, as in the case of insects visiting flowers, but is kept up by the presence of the egg and growing larva during a considerable portion of the period of active vegetable growth, and this has been going on for thousands, probably millions, of years. Yet neither do oaks nor any other plants produce galls spontaneously, as they certainly should do if the results of irritations are in any general sense hereditary. This seems to me to be a really crucial experiment continually repeated by nature.
I may here remark that Mr. Henslow's theory utterly
X 2
breaks down owing to the want of any conceivable connection between insect irritation and most of the innumerable adaptations of the parts of flowers to attract insects and secure cross-fertilisation. Such are the sticky glands, the elastic filaments, the springs and traps, and the accurately timed motions of the pollinia in orchids; the innumerable complexities in papilionaceous flowers; the large coloured bracts in Bougainvillea, Poinsettia, and many others; the flowers with tightly-closed lips, as Linaria, Antirrhinum, Melampyrum, &c.; the enlarged rays of Compositæ, Umbelliferæ, and Caprifoliaceæ; the general massing of small flowers into heads, umbels, corymbs, or dense racemes, so as to become conspicuous, and many other characters. To these may be added the negative evidence of the numerous genera and orders of regular flowers, such as Campanula, Rosaceæ, Gentianaceæ, and many others, which, though thoroughly adapted for insect fertilisation, and whose lower petals have therefore been always subject to irritations, have never developed irregular flowers. In all these cases variation with Natural Selection will account for the phenomena, while insect irritations, even if we admit heredity, will not do so. From whatever point of view we approach the question, the attempt to explain floral structure and colour without the aid of Natural Selection is a hopeless failure.
The Origin of Spines and Prickles.
In the Journal of the Linnean Society ("Botany," No. 208, July 10, 1894) there is an elaborate paper by Mr. Henslow on "The Origin of Plant-Structures by Self-Adaptation to the Environment, exemplified by Desert or Xerophilous Plants," in which the author still further develops his view as to the influence of the direct action of the environment unaided by selection. The only portion of this paper on which I propose to remark is that dealing with the origin of spines and prickles, on which I have already had occasion to write in my book on Darwinism, when combating Professor Geddes' views on the same subject. Mr. Henslow imputes the spines
and prickles of so many plants inhabiting dry countries to the direct influence of the conditions under which they live. This, he thinks, is proved by some of these plants losing their spines when grown under other conditions; he adduces numerous examples of the abundance of spiny plants in such countries as Nubia, Abyssinia, and the Kalahari Desert; and he again and again reiterates the statement that these characters are "simply the inevitable results of the action of environment."
Now if these statements comprised all the facts, that is, if in all dry countries spiny plants abounded, while in all moist or fertile districts they were absent or very rare, the explanation given of their origin would have some plausibility. But there is no such general coincidence of aridity of soil or atmosphere with abundance of spiny plants, as very little inquiry will show. Mr. Henslow points out several other plant-characteristics which indicate, and, as he thinks, are directly caused by, aridity. Such are very small, coriaceous, or rolled up leaves, or their complete absence; a hairy or woolly covering to the whole plant; succulent foliage; special protection of the buds; enormous development of roots; abundance of bulbs and tubers; together with thickness of bark and various protective coatings to stems and leaves. Now many of these peculiarities are present in the flora of the Brazilian Campos—as well described in the memoir of Eug. Warming on Lagoa Santa—which is referred to by Mr. Henslow as corresponding in many respects with that of other arid regions. Yet the author of this memoir expressly states that "spiny plants are very rare" (p. 463). Again, the plants of the Galapagos present similar indications of aridity—shrubs with minute and almost invisible leaves, for example—yet, except the cacti, which may be of American origin, none of the endemic species are spiny. So, also, the rich Sandwich Island flora contains hardly a single endemic spiny plant; and I am informed by the Rev. R. P. Murray, who is well acquainted with the botany of the Canaries, that spiny plants are exceedingly rare in those islands, though much of the surface, owing to the porous volcanic rock and the long periods of drought,
present the conditions which elsewhere are said to produce spines.
Now without denying that—other conditions being equal—aridity may favour and moisture may check the growth of spines, there is another and altogether different set of conditions which seem more directly connected with their abundance or rarity. This is, the presence or absence of herbivorous mammals, against whose ravages spines are a protection. The most destructive of these animals are camels, goats, and antelopes, and it is where these are indigenous—in Arabia, North-east and South Africa, and Central Asia, that thorny shrubs and trees are especially abundant. Again, few countries have more spiny plants than Chili, where the camel-like vicuguas and alpacas, as well as large rodents, are very destructive. But the country is not especially arid, and the remarkable Puyas, whose leaves are armed with excessively sharp recurved spines, inhabit the subalpine regions where rain and mist prevail. In our own moist islands we have a full proportion of prickly plants, and the same may be said of North America, where the Gleditschia or Honey Locust has the young branches, and in old trees the trunk, armed with groups of very strong and sharp spines. So also in Japan, notwithstanding its moist insular climate, we have an Olea and an Osmanthus with holly-like prickly leaves; while the prickly Berberis Darwinii is found in the damp atmosphere of the Straits of Magellan.
Equally opposed to the theory of aridity as the efficient cause of spines is their abundance on palms growing in the hottest and moistest regions of the globe. In many Amazonian species the stem is thickly set with long and very sharp spines pointing downwards, and thus forming a complete protection against monkeys and other arboreal fruit-eating mammals. Many species of Bactris and Astrocaryum are thus armed, as is also the beautiful Guilielma speciosa, the Peach palm, whose fruit is large and edible. It is a suggestive circumstance that, with the exception of palms, few large trees are spiny, and when they are so, as in the case of the Gleditschia, the spines are most abundant on the trunk and on the younger branches. In
the same way, our holly, when it grows to a large size, usually has the leaves towards the top spineless: the wild pear also is spiny below but unarmed above. The climbing palms, on the other hand, are armed to the very top, but in this case the spines assist climbing.
The anomaly of the flora of the Brazilian Campos, having many of the true xerophilous characteristics yet being almost wholly without spiny forms, is quite in harmony with the fact of the great poverty of this region in mammals destructive of woody vegetation. There are really none but a few deer and cavies, which are mostly inhabitants of the more thickly wooded valleys, and which are kept from undue multiplication by the considerable number of species of Felidæ and Canidæ in the same area.
We are, therefore, led to conclude that the apparent direct dependence of an unusually spinescent vegetation on arid conditions of soil or climate is to a great extent deceptive. Such conditions are inimical to the growth of dense forest, and it is a well-known fact that the larger mammalia abound most in partially wooded or open country. Many of these animals are exceedingly destructive to shrubby or arborescent vegetation, especially in districts which are subject to occasional droughts; and it is in such areas that so many of these plants have acquired the protective armature of spines or prickles, while others not so protected have sooner or later succumbed, thus leading to a preponderance of the former. But the numerous instances in which considerable areas and extensive floras are found to have hardly any spinous plants, as compared with other areas in which the soil and climate are generally similar and where such plants abound—the only important difference being the absence or presence of destructive herbivorous or frugivorous mammals—show us clearly that it is the latter rather than the former condition which is the real starting point and efficient cause for the development of spines, while the mode of their production has been through spontaneous variation and Natural Selection.1
1 Professor A. Kerner gives an admirable account of the various forms of spiny and prickly plants, which are exceedingly numerous in the Mediterranean district, and he adds: "In northern regions not exposed to summer drought, where grazing animals find in summer enough green fodder, this form of plant is almost entirely absent." (The Natural History of Plants, English Translation, vol. i., p. 445.)
Adaptations not Explicable on the Theory of Response to Irritation.
A few remarks may now be added on the general question of adaptation in the vegetable kingdom. Reference has already been made to the numerous cases in which the special adaptations of flowers to insect-fertilisation can by no stretch of imagination be imputed to the direct action of insects, and the same thing is equally clear in many other directions. The whole group of insectivorous plants, for instance, exhibit strange and complex adaptations which have no direct relation to the mere fact of insects crawling over them or settling upon them. So also are those varied adaptations by which, as Kerner has shown, injurious insects are prevented from reaching the flowers.
Even more unintelligible on this theory are modifications of fruits and seeds, by which some attract birds or mammals to eat them, while others are guarded against being eaten; some seeds have beautiful wings or plumes for wind dispersal, others have hooks or sticky hairs which cling to wool or feathers, while others again are scattered abroad by the sudden elastic bursting of the capsules. Take the comparatively simple case of nuts. Did they acquire their hard covering and brown protective tints and detachment from the tree as soon as ripe by the direct agency of birds, or monkeys, or squirrels? Of course, the question is absurd, since those eaten by these creatures could not transmit their special qualities; but those that, by the possession of any of these qualities, escaped being eaten, would transmit those qualities to the next generation.1
Any conceivable direct action of the environment can therefore have produced only a very small portion of the modifications and adaptations that actually exist. In by
1 Other cases of the want of relation between adaptations and their supposed cause are given in the next chapter.
far the larger number of cases no such explanation is possible, and no other adequate explanation has been suggested except variation and Natural Selection. It is, of course, admitted that the action of the environment does produce definite changes in all organisms, more especially in plants, but there is no evidence that such changes are transmitted to the offspring of the individuals in which they have been produced.
On the other hand, there is direct evidence that many such changes are not transmitted, an example of which is the Arabis anachoretica with remarkable tissue-papery leaves, due to its growth in hollows of rocks, where neither sun nor rain reach it. Seeds of this plant when cultivated at Kew produced the common Arabis alpina. The same thing occurs with many plants, as every cultivator knows; but other species with no greater peculiarities externally preserve their characters under cultivation, though exposed to the most varied conditions. As we thus know that some variations directly due to the environment are not transmitted, and also know that an immense number of spontaneous or congenital variations are transmitted, since by taking advantage of this fact almost all the improvement in our domestic animals and cultivated plants has been effected; and yet further, that no case has been found in which such spontaneous variations are wholly intransmissible—the logical conclusion is that the two kinds of variation are distinct in their nature. This view of the subject is adopted by those botanists who are now endeavouring to determine the true nature of the numerous alleged species, sub-species, and varieties of our native plants. They test the fixity of the characters which distinguish each form by cultivation. If these characters remain unchanged, and are transmitted by seed, the form is a permanent one and deserves to be recorded as a species or sub-species; but if, as frequently occurs with forms which appear quite as distinct as those which are stable, the plant reverts on cultivation to some other form, it is evidently a modification due to some local conditions of the environment, and should be treated differently. Mr. Beeby has proposed to call the former "intrinsic," the
latter "extrinsic" varieties, terms corresponding to Weismann's "germ variation" and "somatic variation," and these can in many cases only be distinguished from each other by the test of cultivation under different conditions. On this point Mr. Beeby remarks:—"The most transient states of plants due to the direct action of their environment are often far more distinct in appearance from their normal forms than are some varieties from their types; but the first-named return at once to their normal state on being removed from their special surroundings, while the latter remain permanently distinct from their types even when grown under circumstances most disadvantageous to the continuation of the particular variation. That these two kinds of variation exist in plants is certain; and the separation of them seems to be the very basis on which all investigations of the Phanerogamia must be made, if it is hoped that this branch of botany is to throw any further light on "Evolution."1
In conclusion, I submit that the whole body of facts in relation to the direct action of the environment indicates that modifications thus produced in the individual are not transmitted to the offspring; and that until it is demonstrated by experiment that they are so transmitted, theories of plant modification founded on that assumption are altogether worthless.
1 "On the Flora of Shetland." Annals of Scottish Natural History, January, 1892, p. 52.
CHAPTER XVI
ARE INDIVIDUALLY ACQUIRED CHARACTERS INHERITED?
THE question which forms the title of this chapter will not, perhaps, strike the general reader who is unacquainted with recent developments of biology as being of much importance, or as having any special interest for the world at large. Yet it really involves considerations hardly less far-reaching than evolution itself, since the correct answer to it must depend upon, and be a logical consequence of a true theory of heredity. If, then, we can arrive at this correct answer, either by means of observation of natural phenomena or by experiments with living organisms, we shall possess a criterion by which to judge between rival theories; while the answer itself will be found to have a direct bearing of a very important kind on possibilities and methods of human improvement.1
Theories of Galton and Weismann.
Up to about ten years ago the answer to the question would have been almost unanimously in the affirmative. Darwin accepted the inheritance of such characters as an undoubted fact, though he did not attach much importance to it as an agent in evolution; and his theory of pangenesis was an attempt to explain the phenomena of heredity in accordance with it. Mr. Francis Galton made
1 This aspect of the question is discussed in a later chapter.
some experiments which led him to doubt the correctness of Darwin's main contention—that minute gemmules from every cell in the animal body were collected in each of the germ and sperm cells, and thus led to the reproduction of a similar body. He transfused large quantities of blood from black to white rabbits, and vice versa, without in any way injuring them; and after this infusion of blood from a very distinct variety, sometimes to the extent of one-third of its whole amount, each kind bred as true as before, showing no signs whatever of intermixture. He then developed a new theory of heredity, which appeared to him more in accordance with facts, and an essential part of this theory was that the germinal material passes direct from parent to offspring, instead of being produced afresh from the various parts of the body; and, as a consequence, changes produced in the body by external agencies during its life will not be transmitted to the offspring. A few years later Professor Weismann, of Freiburg-in-Baden, independently arrived at a somewhat similar theory, founded on the embryological researches of himself and other biologists; and he supported it by such a body of evidence and by such a wealth of illustration and reasoning that it at once attracted the attention of biologists in every part of the world. This theory being manifestly opposed to the inheritance of acquired characters, he was led to examine the evidence for this dogma, and found it to be extremely scanty, and for the most part quite inconclusive. But as some biologists of great eminence believe that the inheritance of such characters is absolutely necessary in order to explain many of the phenomena of evolution, the discussion on this point has been carried on by many who would have felt little interest in the problem if it were one of embryological development alone. Year by year the question has been discussed in books, pamphlets, and review articles, while Professor Weismann has continued his studies on the whole subject, and in a volume of which an English translation has just appeared, has worked out his theory in very minute detail, grappling fairly with all the various phenomena to be explained, and thus putting the whole
question before the scientific world in a manner which allows it to be fully discussed, tested, and controverted.1
This detailed theory is far too complex and technical to be explained in a short chapter; but as its truth implies that the inheritance of acquired variations is not a law of nature, and as Mr. Herbert Spencer has recently set forth some fresh arguments in favour of such inheritance, and has also reinforced some of his former arguments (in two articles in the Contemporary Review), while an American naturalist has just issued a work,2 in the introduction to which he discusses the same question, and summarises what seem to him the strongest arguments that have been advanced on both sides, concluding also in favour of the inheritance of such characters—a good opportunity is offered to review this evidence, and to show, as the present writer thinks he can show, that all the alleged facts and arguments are inconclusive, and that the balance of the evidence yet adduced is altogether in favour of such characters not being inherited.
What are Acquired Characters?
It is first necessary to understand clearly what is meant by "acquired characters," as even naturalists occasionally miss the essential point, and take any peculiarity that appears in an individual during life to be an "acquired character." But such peculiarities are usually inherited from some ancestor, unless they can be clearly traced to some special conditions to which the individual's body has been exposed. As an illustration, let us suppose twin brothers, very similar in all physical and mental characters, to be subject during life to very different influences: one being brought up from childhood to city life and kept closely at a desk till middle age, the other living always in the country and becoming a working farmer. If the
1 The Germ Plasm: a Theory of Heredity, by August Weismann. Walter Scott. London, 1893.
2 Evolution of the Colours of North-American Land Birds, by Charles A. Keeler, California Academy of Sciences, San Francisco, January, 1893.
one were then pale, slender, weak, and delicate, the other ruddy, stout, and strong, these differences would be, in one or the other, probably partly in both, "acquired characters." And if both, at the same age, married twin sisters, equally alike in childhood but who had been each subject to corresponding conditions in after life, the common idea is that the children of the city couple would be inherently weakly, those of the country couple strong; and that the balance would not be restored even if these two families of children were subject, during their whole lives, to identical conditions. In other words, it is usually believed that the acquired characters of the parents would be transmitted to the constitutions of the children. But it is now asserted, by Weismann and his followers, that facts do not agree with this assumption, and that, in the case supposed, both sets of children would inherit the original qualities of the parents, modified, perhaps, by qualities or characteristics of remoter ancestors, but would not exhibit any effects of the changes produced in their parents by external conditions only.
This latter belief is, I am informed, held and acted upon by breeders of animals as the result of their extensive experience. If a young dog or horse of high breed and good external points becomes accidentally lamed, so as to be permanently disabled from the usual work of its kind, it is often kept for many years to breed from, in full confidence that its offspring will inherit the good qualities of the stock, and will be in no way deteriorated by the absence of work calculated to strengthen the muscles, enlarge the chest, and otherwise increase the power and activity of the parent.
Results that should be Produced by such Inheritance.
Again, if the effects of the use of certain muscles, or of special mental faculties with their corresponding nervous and muscular co-ordinations, were transmitted to offspring, then definite results ought to have been so frequently produced as to have become embodied in general experience and popular sayings. Take the case
of any mechanic working at his trade, whether blacksmith, carpenter, watchmaker, or any other art leading to the use or disuse of special muscles or faculties. If long-continued exercise in one direction leads to increased strength and skill which is passed on to the children, then it ought to be an observable fact that the younger sons should have more strength and skill in their father's business than the firstborn; but, so far as I know, this has never been alleged. So with men of genius, whose mental faculties have been fully exercised in special directions, whether as men of science, artists, musicians, poets, or statesmen; if not only the inherent faculty but also the increased power derived from its exercise be inherited, then we ought frequently to see these faculties continuously increasing during a series of generations, especially through the younger sons, culminating in some star of the first magnitude. But the very reverse of this is notoriously the case. Men of exceptional genius or mental power or mechanical skill appear suddenly, rising far above their immediate ancestors; and they are usually followed by successors who, though sometimes great, rarely equal their parent, whose pre-eminent powers seem generation after generation to dwindle away to obscurity. As illustrations of this principle we may refer to such men as Brindley, Telford, Stephenson, Bramah, Smeaton, Harrison (inventor of the chronometer), Brunel, Dollond, Faraday, Wren, John Hunter, and many others, who were mostly self-taught, and derived nothing apparently either from the faculties or the acquired powers of their parents. So almost all the great poets, musicians, and artists of the world start up suddenly and leave no equals, far less superiors, among their offspring or their descendants. These various classes of facts not only lend no support to the theory of the transmission of acquired faculties from generation to generation, but are not what we should expect if such transmission were a fact. They certainly serve to throw doubt upon it and to show that inheritance is not such a simple matter as this theory implies; they may, therefore, prepare the reader to consider with impartiality the facts and arguments that have been put
forward in its favour, together with the reasons I shall give for the inadequacy of those arguments. For it must always be remembered that the inheritance of this class of characters or qualities must be proved by facts that admit of no other interpretation, if it is to be accepted as one of the bases of the theory of organic evolution. When such tremendous issues are at stake we cannot base our faith upon probabilities, unless they are to an overwhelming degree greater than the probabilities on the other side.
I propose to waste no time on the question whether mutilations are ever inherited, because both parties are now agreed that this is not the point at issue. What we want to know is, whether the effects produced during the lives of individuals by such natural causes as the use or disuse of certain muscles or organs, change of food, or change of climate, are transmitted to offspring, so as to accumulate such effects and thus serve as an important factor in evolution.
Argument from Habits and Instincts.
Two of the cases which have been adduced as affording very strong evidence of the inheritance of an acquired character are, the habit of dogs to turn round several times before lying down, and the peculiar play-habits of the bower-birds; these being supposed to be beyond the power of natural selection to produce, because neither are of vital importance to the species. But such cases as these really prove nothing, because so much in them is hypothetical. It is only guessed that the dog's habit is derived from wild dogs turning round to make a comfortable bed in rough grass. But even if this be a fact, there are many awned seeds of grasses which prick the skin, and in some cases work their way into the body, causing wounds or death, and the turning round may have the effect of laying these awned seeds parallel to the hairs and thus prevent them from penetrating the skin. If so, natural selection would produce and preserve the habit. Again, it may, with many dogs, be a matter of simple
comfort, the turning serving to keep the rather stiff hair all the right way, and also to brush away small hard objects. Yet again, it is not alleged that all dogs do it, and in many cases it may be a habit copied from the mother. The uncertainties of the case are therefore too great for it to afford an argument of any value. The bower-bird's habits are more difficult to explain on any theory, since the whole question of these alleged instincts is unsettled. We have evidence that in many cases even the peculiar song of birds is not instinctive in the species, but is the result of imitation; and Mr. Hudson has recently shown that the fear of man in wild birds or its absence, is probably the result of individual experience in all cases. Till we know that the bower-birds' habit is wholly due to inheritance and not to imitation of older birds, we can hardly found any important conclusions upon it.
Horns and other Dermal Appendages.
Many writers have laid stress on the difficulty of accounting for the origination of new organs in certain groups of animals, by variation and natural selection alone. Horns are especially adduced; and it is alleged that there is no other way of explaining their origin except by the habit of butting with the head, leading to thickening of the skin and excrescences of the bone, which, being transmitted by inheritance and increased by use, gradually produce the various kinds of horns. In like manner, the origin of flowers and their successive modification, have been imputed to the irritation caused by insects, leading to outgrowths which have been inherited and increased by further irritation.
Taking the case of the horns, Mr. J. T. Cunningham, in his introduction to the English translation of Eimer's Organic Evolution, says:—"No other mammals have ever been stated to possess two little symmetrical excrescences on their frontal bones as an occasional variation; what then caused such excrescences to appear in the ancestors of horned ruminants? Butting with the forehead would
VOL. I. Y
produce them, and no other cause can be suggested which would."
This assertion, that "butting with the forehead would produce them," assumes the whole question at issue. There is, I believe, no evidence of it whatever, and there is much that renders it improbable. And the first part of the statement is also erroneous, for Darwin tells us, "In various countries horn-like projections have been observed on the frontal bones of the horse; in one case described by Mr. Percival, they arose about two inches above the orbital processes, and were very like those of a calf from five to six months old, being from half to three quarters of an inch in length." As no known animal in the ancestral line of the horse had horns, these must have been "new characters"; and had they appeared before the ancestral horses acquired such powerful weapons of offence in their hoofs, they would probably have been preserved and increased by selection into formidable weapons. That horns have not unfrequently arisen from such apparently uncaused variations, is indicated by the remarkable difference of structure and growth in the horns of such nearly allied groups as the deer and the antelopes, which at a quite recent epoch must have originated independently. Very suggestive is the curious enlargement of the skull under the crest of the Polish fowls. In another fowl's skull, figured by Darwin, there are two large rounded knobs on the forehead, forming perfect incipient horns.
Dermal appendages, which could not have been caused by special irritations, are so frequent, that almost any useful development appears possible. The spines of the hedgehog and the quills of the porcupine, are of this nature, as are the plates of the armadillo and the scales of the pangolin. The feathers of birds are one of the most marvellous of these developments which, when they once arose, were preserved and modified in endless ways. So, the curious erectile appendage on the forehead of the South American bell-birds, and the equally strange feather-covered cylinder pendent from the throat of the umbrella-birds, are other illustrations of these abnormal outgrowths
of the skin for the origin of which we can assign no cause. Many other secondary sexual appendages of birds are equally inexplicable in their beginnings, such as the long feathers springing from the sides of the head in the six-plumed bird of paradise, and the singular pair of long white feathers growing from among the upper primary wing-coverts of Semioptera wallacei, to which I believe there is nothing similar in the whole class of birds. These various cases of dermal appendages are sufficient to indicate that variations of this kind are continually occurring, which, whenever useful, have been seized upon and increased by natural selection, since any such variations appearing among our domesticated animals are found to be strictly inherited.
Evolution of Teeth.
The American naturalists lay much stress on the evolution of the teeth of mammals in complete palæontological series, alleging that the successive modifications of the cusps conform strictly to lines of use and disuse, and that they are therefore produced by use and disuse. To this there are two distinct replies, either of which seems to me sufficient. In the first place, in such vitally important organs as the teeth of mammals, natural selection will necessarily keep them on these lines, because use implies utility, and disuse, inutility, and utilities necessarily survive. If, then, variations occur in the forms of the cusps—and they certainly do—natural selection will modify them along these lines of utility; and it will be absolutely impossible, from a study of the series of fossil forms, to prove that they have been directly modified by use and that the modifications have been inherited, and that they are not the result of normal variations accumulated by survival of the fittest. The second reply is that of Professor Poulton, who points out that the form of the tooth is fixed before it cuts the gum and that use only wears the cusps away. It is therefore difficult to see how such use in the parent can determine any definite variation in the teeth of the next generation.
Y 2
There is also a general argument, in the fact of so many special types of teeth having been developed, which cannot have been produced by the corresponding use. Such are the arrangement of enamel and dentine in the incisor teeth of rodents, so that they preserve a continual chisel-like cutting edge, and, unlike the teeth of most other mammals, keep on growing at the root so that they are pushed up as fast as they wear away; and the remarkable molar teeth of the elephants, which come forward in succession, and by the arrangement of the folds of hard and soft material always keep a grinding surface, while the enormous tusks grow on continuously during life. These and many other singular modifications of teeth can certainly not be traced to corresponding diversities of use as directly producing them, while they are easily explained by the great variability of all complex organic structures, furnishing material for endless modifications according to the various needs of the widely different mammalian types.
Argument from Tactual Discriminativeness.
We have now to consider Mr. Herbert Spencer's objections in the articles already referred to, which bear the formidable title, "The Inadequacy of Natural Selection." The first of these objections is founded on Weber's experiments on the sense of touch, showing that the power of distinguishing the sensations produced by two points rather close together varies greatly in different parts of the body, the tips of the fingers being able to distinguish the points of compasses when the twelfth of an inch apart, while on the middle of the back they have to be opened more than two inches in order that the pressure of two points may be distinguished from that of one. Between these extremes tactual discrimination varies in different parts of the body, apparently without much relation to utility, except of course in the case of the fingers; and after detailing these at some length Mr. Spencer asks how these divergences can possibly be explained by natural selection. "Why," he asks, "should the thigh near the knee be twice as perceptive as the middle of the thigh?"
And he urges that, in order to prove that these many small variations in different parts of the body have been produced by natural selection, it must be shown that they have influenced self-preservation. He then goes on to show that this perceptive power can be increased by exercise; and concludes with a theory that the differences of tactual perception in different parts are proportioned to the amounts and varieties of contact with substances to which they are subjected, and that these variations in amount and kind of contact have produced diversities of sensibility in the individual, which, by inheritance, have been accumulated in the offspring.
Now, this whole inquiry, and the conclusions drawn from it, seem to me (with all respect for Mr. Spencer's great abilities) to afford a glaring example of taking the unessential in place of the essential, and drawing conclusions from a partial and altogether insufficient survey of the phenomena. For this "tactual discriminativeness," which is alone dealt with by Mr. Spencer, forms the least important, and probably only an incidental portion of the great vital phenomenon of skin-sensitiveness, which is at once the watchman and the shield of the organism against imminent external dangers. The sensations we receive by means of the skin, of contact with various substances, smooth or rough, blunt or pointed, dry or wet, cold or hot, whether indifferent, or pleasurable, or painful, or agonising, afford us information and safeguards without which we could, in a state of nature, hardly exist uninjured for a single day. And we shall find that the delicacy of this absolutely essential danger-signal is almost exactly in proportion to the vital importance of the part to be protected from danger. Thus the maximum of sensitiveness is found in the eye and its surrounding membranes, not because they are most frequently subject to a variety of contacts, for the very reverse is the case, but because this organ is at once the most delicate and the most important for the safety of the individual. So the hands and feet are not very sensitive in those parts which are specially adapted to come in contact with external objects, but in those parts where the tendons, nerves, and blood-vessels which
render them effective organs are most exposed to injury, as in the palm of the hand and the hollow of the foot, and especially on the under side of the arm just above the wrist. The parts of the body which are less sensitive are those where there are masses of muscle, the puncture of which would not cause any serious injury. It will thus be seen that sensitiveness to pain from external agencies is not at all proportioned to the frequency of contacts, but to the vital importance of the parts to be protected; and it is, therefore, such as could not possibly have been produced by inherited use, but must have been developed solely by the preservation of useful variations; and as it is essentially a life-preserving faculty this would inevitably have been effected.
It seems most probable that the faculty of tactual discrimination of adjacent points is partly an incidental result of the distribution of nerve-endings required by skin-sensatien as a preservative faculty, and partly the result of use and attention in the individual. All the facts adduced by Mr. Spencer are in accordance with this view, while none of them in the slightest degree necessitate inheritance of individual experiences to account for them. To show any probability of such inheritance it must be proved that this tactual discriminativeness is a special faculty, due to a different set of nerves from the known nerves of sensation, which, I believe, has never even been suggested. But if it is due to the same nerves, then to separate this small and comparatively unimportant function of skin-sensation from the great and vitally important functions the faculty subserves, and to found upon this artificially isolated and unimportant fragment an argument against the adequacy of natural selection, is not only quite inconclusive, but, as an argument, is altogether null and void.
Argument from Rudimentary Eyes.
The next point discussed by Mr. Spencer is the mode in which the eyes of the Proteus of the Carniola caverns have been reduced to a rudimentary condition; his argument being, that, unless the reduction in size by disuse was
inherited, there is no means of accounting for the almost total disappearance of the eyes. It may be stated that the views held by the followers of Weismann in this country are, first, that as natural selection is always at work to keep all important organs when in use up to their full size and efficiency, the withdrawal of natural selection when the organ ceases to be used, termed by Weismann panmixia, will, by allowing the most imperfect as well as the most perfect eyes to survive, reduce the average size and quality considerably. Then comes the consideration that in total darkness such a delicate organ as the eye would be subject to frequent injury, producing inflammation, and either directly or as a secondary result, death; thus again weeding out those with the largest and most prominent eyes, or those which kept them open habitually in the effort to see. And, lastly, there would come into action what is called economy of growth, the diminution of any useless but complex organ being beneficial, owing to the saving of material and growth-power in building it up.1 It is with this last factor that Mr. Spencer deals, and endeavours to show that the economy would be infinitesimal, because the weight of the eye of this animal is so small. He supposes the reduction to be effected in two thousand years by decrements of one two-hundredth part every ten years; and taking the original eye to have weighed ten grains, he almost laughs to scorn the idea that such an almost infinitesimal amount of diminution at any one time could have given the animals in which it thus diminished a greater chance of survival.
Now, there are two very serious oversights which entirely invalidate this argument. The eye is treated as if it were mere protoplasm weighing so many grains, instead of being a highly complex organ with which muscles, blood-vessels and nerves are connected and co-ordinated in greater proportion perhaps than in any other
1 I have omitted Professor Ray Lankester's suggestion, of a process of selection owing to those individuals with more perfect eyes occasionally finding their way out, because the ancestral immigrants were probably carried far into the caverns by torrential floods, and could only escape by following the water to some of its outlets, success in which would not depend on special acuteness of vision.
organ. I presume the original eye of the ancestral Proteus must have had its three distinct sets of nerves—those of vision, of sensation, and of motion—involving in their normal use the expenditure of a considerable amount of nervous energy, besides the various muscles and blood-vessels connected with it. To measure the benefit to be derived from the entire suppression of such a complex organ, when it became useless, as no more than from the gain of so many grains of simple muscular tissue, appears to me to be an extraordinary misconception of the conditions of the problem.
The second oversight is in ignoring the tremendously severe struggle for existence that would necessarily arise when an animal which had heretofore had the full use of eyes in obtaining food, avoiding danger, and finding its mate, had to enter upon a perfectly new kind of existence, in total darkness, and, moreover, in a place where all kinds of vegetable or animal life were so scanty that the wonder is how those individuals who were first carried into the cavern escaped starvation. Under such conditions as these, would not the various modes of reduction of the eyes above suggested act with an energy and rapidity far beyond their action under normal conditions? And might we not expect the most extreme variations in the direction of the abortion of the eye and of its connected tissues, muscles, and nerves, to have an exceptional value when the food required for building up the organism could only be obtained with the greatest difficulty and in the most limited quantities? Under such conditions I should not be surprised if the greater part of the actual eye-reduction had been effected in fifty or a hundred years, instead of in two thousand. But whether this were the case or no, it will, I think, be admitted that to ignore all these very exceptional conditions, and to argue the case as if the whole matter were one of the economy of a few grains of tissue to an animal whose food-supply was normal, does not add anything to the evidence for the inheritance of acquired variations.
Argument from the Giraffe.
The only other argument of importance adduced by Mr. Spencer is that drawn from the giraffe. This argument was first stated at some length in his essay on The Factors of Organic Evolution, in 1886; and it is now repeated and enforced by some additional considerations, although, Mr. Spencer says, he has met with nothing that can be called a reply; and he adds, that my contention (in Darwinism) that what he alleges cannot be done by natural selection has been done by artificial selection—assumes a parallelism that does not exist. I therefore propose to examine this case more carefully, and shall show that the parallelism I assume is a very close one, and that natural selection is, as Darwin himself believed, fully competent to account for the facts.1
Mr. Spencer's argument is, briefly stated, that to develop such an animal as the giraffe from some antelope-like ancestor requires many coincident and co-ordinate variations of different parts—each increase in the length of the neck, of the head, of the fore or hind limbs, or of either of the bones composing them, requiring corresponding increase of muscles, nerves, and blood-vessels, not only of such as are immediately connected with the enlarged limb, but often in remote parts of the body whose motions are necessarily co-ordinated with it. He maintains that any increase of one part without the adjusted increase of other parts would cause evil rather than good; and that want of co-adaptation, even in a single muscle, would cause fatal results when high speed had to be maintained while escaping from an enemy. Then, again, not only the sizes but the shapes of the bones have to be altered as the muscles are increased in size and the motions of the various parts of the body change; and this introduces fresh difficulties which are, again and again, declared to be insurmountable. And after elaborating all these alleged difficulties at great length, he arrives at the conclusion that, unless the increase or modification of parts
1 Origin of Species, p. 177, and more fully and with admirable force and clearness in Animals and Plants under Domestication, vol. ii. p. 221.
due to use by the individual, is inherited, there can have been no evolution.
Now, I believe, and hope to be able to prove, that these accumulated difficulties are almost wholly imaginary, and arise from a neglect to consider known facts of variation and known methods of adaptation. Mr. Spencer accepts the fact that I and others have laid stress upon, that individual variations are continually occurring in all parts of the organism and in all directions, and that the variations of each part are often independent of each other; but he ignores the equally undoubted fact that certain parts are correlated, and very often do vary simultaneously. The diagrams I have given in my Darwinism show this clearly. A considerable number of parts, as the wings and tail, tarsi and toes of birds, usually vary together, either to the same or to a different amount; but all of them sometimes vary independently, and even in an opposite direction to each other; and such irregular variations evidently afford the very best material for natural selection to work upon, since any kind of variation, either coincident or independent, can be rapidly accumulated. This fact alone does away with half of Mr. Spencer's difficulties.
Another considerable portion of the supposed difficulties is created by assumptions which pervade his articles but which are opposed to the facts of nature. He tacitly assumes that natural selection works by the preservation of large individual variations—"sports" as they are often termed; whereas both Darwin himself and all his followers entirely reject these as causes of modification of species (except perhaps in rare cases where they may initiate new organs), and rely wholly on those individual variations which occur by thousands and tens of thousands in every generation. Mr. Spencer continually uses such expressions as, "This one has unusual agility"; "that one develops longer hair in winter"; "another has a skin less irritated by flies"; "it is needful that the individual in which it occurs shall have more descendants"; "a variation… might sensibly profit the individual in which it occurred"; "would an individual… survive"; "favourable variations… would disappear again long before one or a few of the co-operative parts could be appro-
priately varied." The same assumption pervades the writings of most of the opponents of Weismann. Thus Mr. Keeler, in the work already quoted, says, referring to the modification of organs by variation and selection only: "This explanation is open to two objections: first, the one already raised by Mr. Spencer, that before the second correlative variation appeared the first would be lost; and, second, the suggestion of Cunningham in regard to assuming that use could develop the character required, but that the individual thus favoured could not transmit the variation, but that posterity must wait for the same variation to arise spontaneously. This hypothesis is so forced, illogical, and absurd, that so long as a better one can be found it should be adopted."
Now all this implies that there are but few variations occurring at long intervals; the facts being that, in populous species, every generation affords many thousands of variations of every observable part and organ; whence it follows that the individuals of every such species can be divided into two sets as regards each organ or each group of parts, such as those with a longer and those with a shorter neck, those with neck and legs longer and those with the same parts shorter. In the latter case perhaps a quarter or an eighth only of the whole population would be found in each category; but as, in most cases, not one eighth part of those born each year can survive, this would be ample. It will be seen that the facilities for modification are thus indefinitely greater than the expressions and arguments of Mr. Spencer and his supporters assume them to be.
Another tacit assumption is, that in nature all the individuals of a species have their parts so perfectly co-ordinated that any increase of one part only would disturb the harmonious adjustment and be a disadvantage. But this is totally to misconceive the situation. The adjustment of parts is a rough working adjustment, sufficient for the purpose of maintaining life, but capable of being improved (or deteriorated) by very many slight modifications of single parts. To illustrate this general adjustment, let us suppose we have before us for comparison all
the county elevens of English cricketers. We shall have a body of some hundreds of picked men, all of whom are probably above the average as runners, are exceptionally quick with eye and hand, and are all more or less active and muscular. They vary, of course, in their special capacities, whether as batters, bowlers, fielders, or wicket-keepers, but it is certain that most of them would take a high place in almost any form of athleticism to which they chose to apply themselves. Yet these men would not resemble each other closely in stature or proportions. We should find among them tall and short, slender and stout; and among those of the same height proportions would differ, some being long-legged, others short-legged, and perhaps no two of the lot would be found to have exactly the same proportions in all measurable parts of the body. We are thus shown that a high average result of strength and activity can be reached by very various combinations of the bones and muscles of the limbs and other variable parts, and we can hardly doubt that almost all of these men could be rendered still more efficient cricketers or athletes by some slight improvement in their organisation. One would run better if his legs were longer, another would throw and bowl better if his arms were shorter and more muscular; and such changes would be effective because these parts are now imperfectly co-ordinated with the rest of the body.
The considerations suggested by this illustrative case immensely increase the facilities for the improvement of any faculty required by natural selection, and they enable us to understand the process by which both natural and artificial selection have been able to modify the form and qualities of so many animals. It is not, as Mr. Spencer's argument assumes, by the selection of improvements in any special bone, or muscle, or limb that these modifications have been effected, but by the selection of the capacities or qualities resulting from the infinitely varied combination of variations that are always occurring. Horses have been improved by preserving the swiftest for racing, the strongest for the plough, and a combination of speed, endurance, and jumping for the hunting field. Improved
combinations do occur in all these classes in every fresh generation; and breeders know that from, say a hundred half-wild horses, whether from the prairies, the pampas, or the Australian bush, careful selection could obtain—probably in less than a century—very excellent representatives of each of the three above mentioned types of horses.
Recurring now to the case of the giraffe, whose whole organisation has been modified so as to obtain an almost unlimited feeding-ground from bushes and trees far above the limits eaten off by the tallest antelopes and buffaloes, we see that numerous successive modifications have all worked in this one direction—the large size, the enormous shoulders giving a sloping back, the correspondingly long neck, the long head, and the elongated prehensile tongue, so that a full-grown male giraffe can feed up to twenty feet above the ground.
In considering how this was brought about we must remember that the struggle for existence is very intermittent in character. A severe drought causing a scarcity of herbage, and leading the antelopes and buffaloes to compete with the giraffes for the foliage near the ground, may occur at intervals of from twenty years to a century. When such droughts occur, animals that can feed half a foot higher than others would survive; not one individual here and there, but the tenth or twentieth part of the whole population in a given district, amounting, perhaps, to many thousands of individuals. These taller survivors would go on increasing for another long period, in the meantime being subject to the usual struggle with wild beasts and other dangers, natural selection keeping the whole organism up to the mark, till another period of scarcity led again to an elimination of the shortest. Those that survived would not be all alike; some would gain the increased feeding power by higher shoulders, some by longer neck, or longer head, or more extensible tongue, or by various combinations of these variations; and the regular elimination of the weaker or less active in every year of the intervening periods, together with the equalising result of constant intercrossing, would produce a race possessing the one essential character of lofty
feeding power by means of the altogether unique combination of characters found in the giraffe. There has been ample time for the process, probably the whole of the pliocene period; and in view of the known amount of variability in all parts of the organism, and the intermittent nature of the struggle for existence, there seems far less difficulty in the production of this animal than in the production of the beautiful and perfectly co-ordinated greyhound in an almost infinitely shorter time.
And now we see how very close is the parallelism between artificial selection and natural selection, and how purely imaginary are the difficulties of co-ordination set forth so elaborately by Mr. Spencer. For, by the preference of some men for swiftness in their dogs, of others for those which followed game by sight rather than by scent, of others again for elegant slenderness of form, the greyhound, almost as perfect as we now see it, had been already developed in the time of the ancient Egyptians from some wolf-like wild ancestor. Men have always preserved qualities, not single characters, just as nature preserves the qualities of speed, strength, agility, acute scent or vision, not any particular variation of bone, muscle, or member. The two modes of selection are thus strictly analogous and strictly comparable; and the whole elaborate structure of "insurmountable difficulty" founded by Mr. Spencer on the supposed impossibility of adjustment of parts by variation and selection, falls to the ground. As a matter of fact, there is a sufficiency of useful variation always present in each succeeding generation to increase any required life-preserving quality, all theoretical objections to the contrary notwithstanding.
The Small Jaw in Civilised Man.
In his Principles of Biology, Mr. Spencer discussed, briefly, the smaller jaws of civilised races as only to be explained by the inheritance of successive reductions produced by the use of softer food, since any advantage to be gained by each step of the process would be too minute to have any appreciable effect in saving life under any possi-
ble circumstances. In his Factors of Organic Evolution he recurs to the subject, in order to exclude two imaginable causes which he had not before referred to—a correlation of decreased jaw with increased brain, which he rejects, because many small-brained people are also small-jawed, while others distinguished for their mental power have yet jaws above the average size: and sexual selection, which he also rejects for obvious and valid reasons.
Subsequently, the same question was discussed by Mr. W. Platt Ball, in his excellent little work, Are the Effects of Use and Disuse Inherited? He shows that the difference of size is less than has been supposed, while the great range of variation in the size and weight of the jaw, both in civilized and savage races, renders all comparisons useless unless founded on a very large number of specimens. He then accounts for the reduction that actually exists by a variety of suggested causes, some of which are undoubtedly open to criticism. He also states that he "allows for a reduction proportional to that shown in the rest of the skull," because some other agency than disuse must have reduced the thickness of the skull, and, presumably, of the jaws also; after which allowance there remains but a small further reduction of the jaws to be accounted for.
This portion of Mr. Platt Ball's work called forth a very minute criticism from Mr. F. Howard Collins, which, being declined by the editor of Nature—as the author tells us in his preface—has been published in pamphlet form. As Mr. Collins is the author of An Epitome of the Synthetic Philosophy, so well done as to have received the approval of Mr. Spencer himself, he may be supposed to speak with some authority when he undertakes to attack one of Mr. Spencer's critics; and for this reason I think it advisable to point out the erroneous and illogical principles on which his whole argument is founded. In order to compare the jaws of Australians with those of recent Englishmen, he takes one linear dimension of the jaws, cubed to get the proportionate bulk, and compares this with the cubical contents of the skulls to which the jaws belong, that is, with the mass of the brain. Now, as the Australians have very much smaller brains than Europeans, to compare the jaw with the skull-capacity is to make it appear much larger
in the Australian and smaller in the European, even when the actual size is identical. Why should the jaw be measured by the brain as a standard, any more than the foot, the hand, or the stature? On the same principle the Australians might be proved to have very large hands, and to be much taller that Englishmen, whereas, like most savages, they have rather small hands, and are hardly our equals in stature. The only reasonable way of comparing the jaws of two races so nearly equal in stature as Australians and Englishmen, is to compare them directly. The jaw is really a limb, used in the mastication of food to supply the whole body with nourishment, not the brain only; and its true organic relation is with the body, not with the brain case to which it is attached, still less with the brain itself. By the illogical process he uses, of first increasing the linear dimension of the Australian's jaw in the same proportion as the English skull is larger than the Australian skull, without attempting to ascertain whether the depth and thickness increase in the same proportion, and then cubing this dimension, he arrives at the amazing result that the Australian's jaw is very nearly double the bulk of the modern English jaw! Mr. Platt Ball, on the other hand, finds the weight of the latter to be only 5 per cent. less, when reduced in proportion to the lighter skull, the actual difference being 17 per cent.; but both results are founded on far too small a number of specimens to be in any way trustworthy. Mr. Collins then replies, and I think forcibly, to two of Mr. Ball's suggested causes of the decrease—lightness of structure facilitating agility, and sexual selection—but he passes over the two which have most weight, the one as a "tentative suggestion," the other altogether unnoticed; namely, cessation of selection, and increase or decrease caused by use and disuse in the individual. The first, Mr. Collins says, with a strange misapprehension of the point, would only affect the weight and thickness of the jaw, not the peripheral measurement which he has used. But why should not the size—the length and depth—of the jaw be quite as important as the thickness and strength? The greater length and depth of the jaw would be effective in giving more room for the attachment of the muscles on which the whole efficiency of the masticating organ depends
as well as by affording space for the full development of large and well-formed teeth. In the early stages of human progress, when much indigestible food had to be eaten, large bones to be gnawed, and meals to be hastily devoured, the large and powerful jaw would be preserved by natural selection. But civilised man has no need for such a bulky apparatus, hence the average size of the jaw would fall "from the birth-mean to the survival-mean," to use Mr. Lloyd Morgan's neat expression—small-jawed men being at no disadvantage in the struggle for existence; whence the occurrence among us of very small and very large jaws, though with a lower average than among the Australians.
The other point—and I believe a very important one—is the diminution, in the individual, due to comparative use and disuse. From the time the first teeth are obtained, the jaws are used in mastication many times a day, and the difference in the amount of exertion and strain on the muscles and bone, in the case of a civilised European living mostly on soft or well-cooked food and the savage chewing up tough roots and tearing half-raw flesh from the bones of almost any animal he can kill, must be very great. This difference acting while the bones of the face are growing, in the period between childhood and manhood, and to a less degree, perhaps, on to middle age, would certainly lead to a difference of size—and probably to a large portion of the difference that actually exists—between the jaws of savage and civilised man. If we consider, further, that concurrently with the diminished use of the jaws there was an increased use and development of the brain, it may well be that the process of reduction of the former was facilitated by the diversion of a portion of the supply of arterial blood to the latter.
These two causes—cessation of selection or panmixia, and the effect on the individual of greater or less exercise of the parts—are admittedly real causes, their effects can be roughly estimated, and they seem fully adequate to account for the comparatively small difference that actually exists between the jaws of the lowest and highest races now on the globe.
VOL. I. Z
Individual Modification.
The question of purely individual modification by use or environment requires further consideration, because, owing to the unhesitating acceptance of the inheritance of acquired characters by Darwin and most contemporary naturalists, the important bearing of facts proving the effect of external conditions on individuals has been, and still is, altogether overlooked. Two cases in particular are continually quoted by the advocates of inheritance, as if they were in some way antagonistic to Weismann's theory, whereas they really support that theory, and almost prove it. The first is that of the Texan species of Saturnia (Emperor-moth) which feeds on the black walnut (Juglans nigra), and which, when pupæ were brought to Switzerland and the larvæ raised from the eggs laid by the moth were fed on the common walnut (Juglans regia), produced moths which differed so much from the parent both in colour and form that it appeared to be a new species. Darwin, writing to Wagner of this case, says: "When I wrote the Origin, and for some years afterwards, I could find little good evidence of the direct action of the environment; now there is a large body of evidence, and your case of the Saturnia is one of the most remarkable of which I have heard." In referring to this letter, Professor H. F. Osborn, in one of the most intelligent discussions of this question I have yet seen from an American author, says, "Darwin distinctly abandoned the utility principle in the case of Saturnia";1 and Mr. D. G. Elliott, in his presidential address to the American Ornithologists' Union in 1891, quoted the same case as affording striking evidence of the transmission of acquired modifications.
Professor Lloyd Morgan (in his Animal Life and Intelligence, pp. 163—166) sees clearly that this and other cases do not prove more than a modification of the individual; but it seems to me to go further than this.
1 Are Acquired Variations Inherited? An Argument, by Henry Fairfield Osborn, in The American Naturalist, February, 1891.
For here we have a species, the larvæ of which for thousands, perhaps millions of generations, have fed upon one species of plant, and the perfect insect has a definite set of characters. But when the larvæ are fed on a distinct but allied species of plant, the resulting perfect insect differs both in colouration and form. We may conclude from this fact, that some portion of the characters of the Texan species were dependent on the native food-plant, Juglans nigra, and that this portion changed under the influence of the new food-plant. Yet the influence of the native food had been acting uninterruptedly for unknown ages. Why then had the resulting characters not become fixed and hereditary? The obvious conclusion is, that, being a change produced in the body only by the environment, it is not hereditary, no matter for how many generations the agent continues at work; in Weismann's phraseology, it is a somatic variation, not a germ-variation.
The other case is that of the brackish water shrimp, Artemia salina, which, by the water becoming gradually Salter, was changed into what had before been considered a distinct species, A. milhausenii; and the reverse change was also effected, the modification in both cases being in proportion to the alteration in the salinity of the water, and therefore spread over two or three generations. Here too there seems to be no heredity, however long either form has been submitted to the influence of the modifying medium.
Similar to this is the case given by Mr. Thiselton Dyer (Nature, vol. xliii., p. 581) of the tissue-papery leaved Arabis anachoretica, which grows in hollows of rocks sheltered from sun and rain, but which, when cultivated at Kew, changed at once to the common Arabis alpina, of which it is a modification due to the action of the environment on the individual plant. How different is this from the behaviour of plants which have been developed by germ-variation and selection, such as most of the true alpine plants which retain their compact dwarf foliage and large flowers under very different conditions when raised from seed in our gardens! Other
Z 2
species, however, which have a considerable range and become dwarfed in some localities by adverse conditions of drought, wind, or other causes, at once grow to a larger size when cultivated and sheltered. Many of our British botanists are now applying this test to distinguish those forms of our native plants which owe their peculiarities to germ-variation from those which have been modified individually by the action of the environment. We are often unable to decide by mere observation to which class any particular variety or local form belongs, but cultivation at once determines the point; for while the former transmit their peculiarities, however minute these may be, to their offspring, the latter revert at once to the parent form. Mr. Beeby has proposed to call the former class "intrinsic," the latter "extrinsic" varieties, useful terms which indicate that the one are due to an internal cause, are therefore stable, and show us the incipient stage of species-formation; while the other is merely an external modification of the individual which has no stability, being wholly due to the direct action of the environment. It is very important to note the sharp distinction between these two kinds of varieties, externally so alike, though having a fundamentally different origin. There appears to be no gradation from one to the other. The individually acquired or extrinsic character, however long it may have persisted, disappears instantly when the special environment that produced it is changed, as in the case of the Texan Saturnia, the papery-leaved Arabis, and many similar cases; while intrinsic characters—those due to germ-variation—however slight they may be, as in the various races of mankind, many of the closely allied species of moths, and some of the sub-species or varieties of our native plants, preserve their characteristic features under greatly changed conditions.
The cases now given of change in the individual due to external causes and often of a very marked character, render it exceedingly probable that a large portion of the observable difference in the size of the jaws of civilised man and of some domesticated animals, as well as all those changes produced more or less suddenly by a change in
the environment, are mere individual effects which are not hereditary; while, whenever such changes appear in species that have long been subjected to uniform conditions —as in the case of the Texan Saturnia—they indicate that some portion of the external peculiarities of those species were individually acquired characters, and therefore afford strong evidence of the non-inheritance of such characters.
Summary of the Argument.
I have now fairly met, so far as the space at my disposal will allow, the strongest arguments of the advocates of use-inheritance as a law of nature and as a factor in evolution. I have shown that the effects which it ought to produce in the case of mankind do not appear, and that breeders of animals do not recognise it as a factor to be taken account of. The alleged cases of inherited habits or instinct, supposed to be useless, are shown in one case to be not necessarily so; while all such cases involve so many elements of uncertainty or ignorance that no conclusion of value can be drawn from them. The alleged difficulty of the origin of horns except by the inherited effect of blows and pressures, I have shown to be founded on error as to fact; and their origin by normal variation and their development where useful by selection, to be supported by the frequent occurrence of dermal excrescences in many animals. The case of the mammalian teeth has been shown to be quite explicable without use-inheritance, the mode of action of which is, in this case, itself inexplicable. Mr. Herbert Spencer's three main arguments to prove the inadequacy of natural selection have been fully discussed, and have, I venture to think, been shown to be entirely inconclusive, since they are either founded on comparatively unimportant and adventitious peculiarities, or on a neglect of some of the most important conditions under which natural selection in its various forms comes into play.
In order to render the position clearer I wish, before concluding, to say a few words on the general question.
Those who are termed Neo-Darwinians do not yet maintain that use-inheritance (to use Mr. Ball's convenient term) does not exist, but merely that it has not been proved to exist. Whether it actually occurs—and if it occurs at all, I believe it must occur constantly—can only be proved either by very careful and long-continued experiment, or by the demonstration of some theory of heredity which either necessarily includes or excludes it. But even if it does exist I myself believe that it is altogether unimportant as a factor of evolution, and that we have evidence sufficient to prove that natural selection is not inadequate for want of it.
When we urge that the effects of use-inheritance, if it exists, ought to be abundantly visible in some such ways as I have suggested in the early portion of this chapter, its advocates reply, that only a small portion of what is acquired by the parent is transmitted to the offspring, and that its effects may, therefore, only become visible after a long series of generations; but as it is necessarily cumulative it must produce a considerable result in the course of ages. All we can say in reply to this is, that it is pure hypothesis, and that if true, it may serve to explain the difficulty of obtaining evidence in its favour, but at the same time it indicates that use-inheritance can be of no value as a factor in evolution. Variation is so large and so constant that any required character can be greatly modified in a very short time, of which Darwin gives many illustrations. The comb of the Spanish cock was made upright, the comb and wattles of the Polish fowl were completely abolished, and the average weight of ducks was raised from four pounds to six pounds in a few years. Our sheep, pigs, and cattle were wonderfully improved, and often completely changed in form between the latter portion of the eighteenth and the first half of the present century, while many of our flowers and fruits have been nearly doubled in size, and greatly improved in form and colour in the same period. But natural selection has a great advantage over artificial selection in the enormous scale on which it works, giving much greater scope for the occurrence of favourable variations in large numbers.
Thus, whenever some great change of conditions led to a more severe struggle for existence, the modifications of structure needed for adaptation to the new environment would soon be effected without any aid from use-inheritance.
That this is the fact is further indicated by the large range of characters and adaptations that must have been produced by variation and selection alone, since use-inheritance cannot possibly have had any part in their development. Mr. Spencer admits that there are many such, but does not recognise the weight of the argument which they afford against the need of use-inheritance as a factor in evolution. It is well, therefore, briefly to enumerate some of the more important of them. We have already referred to the teeth, in their numerous peculiarities of form, structure, and mode of growth, many of which are quite removed from any direct influence of use in their production. Still less can we impute the hair to such causes, in its varieties of length, thickness, texture, and colour, with its occasional modification into protective armour, such as plates, scales, or spines, or into offensive weapons, such as the horns of the rhinoceros. The bills of birds, in such strange modifications as are presented in the duck, the spoonbill, the heron's spear, the woodpecker's chisel, the snipe's sensitive borer, the enormous but very light bill of the toucan, the powerful nut-cracking bill of the cockatoo, and many others, all evidently adapted to special uses, but by no possibility developed by those uses; the wonderful modifications of the stomach in ruminants, and especially in the camels; the whole series of protective, warning, and recognition colours of animals; the numerous peculiarities of structure and instinct in neuter insects where use-inheritance is absolutely excluded; and, lastly, the whole of the wonderful protective and distributive contrivances of fruits and seeds, and the still more wonderful and more complex adaptations of flowers to insect fertilisation, this latter, be it remembered, not under the pressure of an individual struggle for life, but only for the purpose of obtaining an increase of vigour and a somewhat more rapid multiplication, giving
an advantage over other species in the general struggle for existence.
Even more removed from possible development through use are the special organs of sense, such as the eye and the ear. The complexity of structure in the internal ear is amazing, with its spiral and semi-circular canals and tubes, its membranes, ducts, and cartilages, its extraordinary stirrup, hammer, and anvil bones, its wonderful rods, hair-cells, and otoliths. It seems like some strange machine, the connection of whose various parts and their mode of action it is impossible to follow. Yet we feel sure that every detail has its use; and we have here an organ the co-adaptation of whose several constituent parts is essential to its utility, and is apparently more difficult to bring about by variation and selection than in the cases where Mr. Spencer thinks it absolutely necessary to call in the aid of use-inheritance. But in this case it is utterly unimaginable that any amount of air-waves impinging on the tympanum can have tended directly to the production of this highly complex and delicately adjusted organ. The case of the ear alone appears to me sufficient to prove that use-inheritance is not required for the development and progressive modification of the most complex and beautifully adjusted structures.
We thus see that there is a wide range of characters and structures, often involving the most beautiful adaptations, in which use-inheritance can admittedly have no share; that even if it exists in other cases it is unnecessary, since it can only give a little help in a process which is demonstrably within the power of variation and natural selection; and that the strongest arguments that have been urged, either to show its supposed necessity or to prove its actual existence, break down on close examination, and in some cases even afford strong evidence against it. Our conclusion, therefore, is, that no case has yet been made out for the inheritance of individually acquired characters, and that variation and natural selection are fully adequate to account for those various modifications of organisms which have been supposed to be beyond their power.
CHAPTER XVII
THE METHOD OF ORGANIC EVOLUTION
THE modern doctrine of organic evolution may be said to date from the great French naturalist Buffon, who, more than a hundred years before the publication of the Origin of Species, clearly indicated his belief in the mutability of specific and generic forms, although, owing to the power of the Church in his day, he was often obliged to veil his opinions under the guise of hypotheses, which, as they were opposed to religion of course could not be true. Yet he occasionally speaks very plainly, as when he says:—
"Nature, I maintain, is in a state of continual flux and movement;"
and again—
"What cannot Nature effect with such means at her disposal? She can do all except either create matter or destroy it. These two extremes of power the Deity has reserved for Himself only; creation and destruction are the attributes of His omnipotence. To alter and undo, to develop and renew—these are powers which He has handed over to the charge of Nature."
Dr. Erasmus Darwin held similar views, which he developed at great length, and in doing so, anticipated many of the arguments afterwards elaborated by the celebrated Lamarck, that changes in species were caused both by the direct action of the environment, by the use and exercise by animals of their several organs, and more especially by
the effects of effort and desire leading to the development of parts and organs calculated to gratify those desires. The great French naturalists Geoffrey and Isidore St. Hildaire adopted these views with certain modifications, as did a limited number of German naturalists; while they were popularly set forth with much knowledge and literary skill by the late Robert Chambers in his Vestiges of Creation. Somewhat later the general theory of evolution was explained and illustrated by Herbert Spencer with so much power and completeness as to compel its acceptance by most thinkers; but neither he, nor any of the great writers who had gone before him, had been able to overcome the difficulty of explaining the process of organic evolution, since no one had been able to show how the wonderful and complex adaptations of living things to their environment could have been produced by means of known laws and through causes proved to exist and to be of sufficient potency. Alike for naturalists, for men of science in general, and for students in philosophy, the method of organic evolution remained an insoluble problem.
Darwin himself, who after his return from the Beagle voyage was in close intimacy with the chief naturalists of the day, tells us in his letters of the general feeling with regard to all theories of the evolution of species. In 1844 he wrote to Dr. Hooker—" At last, gleams of light have come, and I am almost convinced, that species are not (it is like confessing a murder) immutable." And again in 1845 to L. Jenyns—" A long searching amongst agricultural and horticultural books and people makes me believe (I well know how absurdly presumptuous this must appear) that I see the way in which new varieties become exquisitely adapted to the external conditions of life and to other surrounding beings. I am a bold man to lay myself open to being thought a complete fool, and a most deliberate one."
Considering that this state of opinion prevailed up to the very date of publication of the Origin of Species, the effect produced by that work was certainly marvellous. A considerable body of the more thoughtful naturalists at once accepted it as affording, if not a complete solution,
yet a provisional theory, founded upon incontrovertible facts of nature, demonstrating a true cause for specific modification, and affording a satisfactory explanation of those countless phenomena of adaptation which every preceding theory had been powerless to explain. Further consideration and discussion only increased the reputation of the author and the influence of his work, which was greatly enhanced by his Animals and Plants under Domestication, published nine years later; and when this had been fully considered—about twelve years after the publication of the Origin—a large proportion of naturalists in every part of the world, including many of the most eminent, had accepted Darwin's views, and acknowledged that his theory of Natural Selection constituted—to use his own words—"the main but not the exclusive means of modification." The effect of Darwin's work can only be compared with that of Newton's Principia. Both writers defined and clearly demonstrated a hitherto unrecognised law of nature, and both were able to apply the law to the explanation of phenomena and the solution of problems which had baffled all previous philosophers.
Of late years, however, there has arisen a reaction against Darwin's theory as affording a satisfactory explanation of organic evolution. In America, especially, the theories of Lamarck are being resuscitated as of equal validity with natural selection; while in this country, besides a considerable number of Lamarckians, some influential writers are introducing the conception of there being definite positions of organic stability, quite independent of utility and therefore of natural selection; and that those positions are often reached by discontinuous variation, that is, by spurts or sudden leaps of considerable amount, which are thus "competent to mould races without any help whatever from the process of selection, whether natural or sexual."1 These views have been recently advocated in an important work on variation,2
1 Discontinuity in Evolution. By Francis Galton. Mind, vol. iii., p. 367.
2 Materials for the Study of Variation, treated with especial regard to Discontinuity in the Origin of Species. By William Bateson, M.A. 1894 (pp. xv. and 598).
which seems likely to have much influence among certain classes of naturalists; and it is because I believe such views to be wholly erroneous and to constitute a backward step in the study of evolution that I take this opportunity of setting forth the reasons for my adverse opinion in a manner likely to attract the attention not only of naturalists but of all thinkers who are interested in these problems.
What the Darwinian Theory really Is.
Before proceeding to this special discussion it may be well to illustrate briefly the essential difference between the theories of Darwin and those of his predecessors and opponents, by a few examples of those cases of adaptation which are insoluble by all other theories, but of which natural selection gives an intelligible explanation.
The Darwinian theory is based on certain facts of nature which, though long known to naturalists, were not understood in their relations to each other and to evolution. These facts are: variation, rapid multiplication, and the resulting struggle for existence, and survival of the fittest. Variation is the fundamental fact, and its extent, its diversity, and its importance are only now becoming fully recognised. Observation shows that when large numbers of individuals of common species are compared there is a considerable amount of variability in size, form, colour, in number of repeated parts and other characters. Further, that each separate part or organ which has been thus compared varies, so that it may be safely asserted that there is no part or organ of animals or plants that is not subject to continual variation. Again, all these variations are of considerable amount—not minute, or infinitesimal, or even small, as they are constantly asserted to be. And, lastly, the parts and organs of each individual vary greatly among themselves, so that each separate character, though sometimes varying in correlation with other characters, yet possesses a considerable amount of independent variability. The amount of the observed variation is so great that in fifty or a hundred adult individuals of the same sex, collected at the same time and place, the difference
of the extreme from the mean value of any organ or part is usually from one-tenth to one-fourth, sometimes as great as one-third of the mean value, with usually a perfect gradation of intervening values.
The multiplication of individuals of all species is so great and so rapid that only a small proportion of those born each year can possibly survive; hence a struggle for existence necessarily follows, the result of which is that, on the average, those individuals which are in any way ill-fitted for the conditions of existence die, while those better fitted live. The struggle is of varied character and intensity—either with the forces of nature, as cold, drought, storms, floods, snow, &c.; with other creatures, in order to escape being devoured, or to obtain food, whether for themselves or for their offspring; or with their own race in the competition for mates and for the means of existence; while as regards all these forms of struggle mental and social qualities are often as important as mere physical perfection, and sometimes much more important. The fact already stated, of the large amount of variability in most species, has been thought by some to show either that there can be no such severe struggle as has been suggested, or that the characters which vary so much can be of little importance to the species, and cannot therefore determine survival. But in making this objection two considerations have been overlooked. In the first place we always compare adults, and an enormous amount of destruction has already taken place during the earlier stages of life. The adults, therefore, are already a selected group. In the second place, the struggle is very largely intermittent, owing both to the occurrence only at long intervals of the most adverse meteorological conditions, while the diversity of these conditions leads in each case to the selection of a different characteristic. An exceptionally severe winter will destroy all which are deficient in one set of characters, while a long drought, or scarcity of some particular kind of food, will weed out those deficient in another set of characters. Thus, in any one year there will exist numbers of individuals which are doomed to speedy de-
struction under some one of the special adverse conditions which are constantly recurring; and it is this, probably, that explains why there is so much individual variation continually present, although the central or typical form remains unchanged for very long periods. This typical form is that which, under existing conditions, survives all the periodical or secular adverse changes, during which the outlying, or extreme variations of whatever kind, are sooner or later eliminated. It is for want of giving full weight to the essentially intermittent nature of the struggle for existence that so many writers fail to grasp its full significance, and continually set forth objections and difficulties which have no real importance.
Darwinism as compared with Lamarckism.
We are now in a position to estimate the efficiency of Darwin's theory in explaining the wondrous and complex adaptations that abound in the organic world, as compared with that of Lamarck or of his modern supporters. And first let us take the simple case of the adaptation of fleshy and juicy fruits to be eaten by birds, causing what seems at first sight an injury to the species, but which is really most beneficial, inasmuch as it leads to the wide dispersal of the seeds, and greatly aids in the perpetuation of the plants which produce such fruits. To what possible direct action of the environment can we impute the production of fleshy or juicy pulp, with attractive colour, and with small, hard-coated seeds, in the innumerable fruits which are devoured by birds, through whose bodies the seeds pass in a state fitted for germination? There is here a combination of characters calculated to a certain end—a definite adaptation. If we suppose that in an early stage of development ancestral fruits which happened to be a little softer than others were eaten by birds, how could that circumstance increase the softness, develop juice, and produce colour in future generations of the trees or bushes that sprang from the seeds so dispersed? And if we assume that these several characteristics are positions of "organic stability," acquired through acci-
dental variation, we have to ask why the several kinds of variation occurred together, or why neither of them occurred in the numerous species in which to be eaten would be injurious instead of beneficial?
But if we begin at the same stage and apply the Darwinian theory we find that the whole process is easy of explanation. It is an observed fact that fruits vary in softness, juicyness, and colour, and seeds in the hardness or hairyness of their integuments. Any variation of primitive fruits in either of these directions would therefore be beneficial, by attracting birds to eat them and so disperse the seeds that they might reach suitable stations for development and growth. Such favourable variations would therefore be preserved, while the less favourable perished.
Now ask the same questions as to the production of the innumerable modes of dispersal of seeds by the wind, from the simple compressed form and dilated margins of many small seeds, to the winged seeds of the ash and maple, and the wonderful feathery parachute of the thistle and the dandelion. Or again, inquire as to the wonderful springed-fruits which burst so as to scatter the small seeds, as in some of the balsams; or yet again, as to the sticky glands of the sundews, and the small water-traps of the bladder-wort; and a hundred other equally strange adaptations to some purpose of use to the species, but whose development has no relation whatever to any possible direct action of the environment, though all of them are explicable as the result of the successive preservation of such variations as are known to occur, acting at various intervals and by means of successive modifications, during the whole period of the development of the group from some remote ancestral form.
The modern advocates of Lamarckism content themselves with such simple cases as the strengthening or enlarging of organs by use, the hardening of the sole of the foot by pressure, or the enlarging of the stomach by the necessity for eating large quantities of less nutritious food. These, and many other similar modifications, may doubtless be explained by the direct action of conditions,
if we admit that the change thus produced in the individual is transmitted to the offspring. That such changes are transmitted has, however, not yet been proved; and a considerable body of naturalists reject such transmission as improbable in itself, and at all events as not to be assumed without full and sufficient proof. But even if accepted it will not help us to explain the very great number of important adaptations which, like those already referred to, are quite unrelated to any direct action of the environment. Having thus cleared away some preliminary misconceptions, and stated in briefest outline the main features of the law of natural selection, we may proceed to consider the objections of those modern writers to whose works we have already referred.
The Theory of Discontinuity.
Mr. Bateson's large and important volume consists mainly of an extensive collection of cases of variation of a particular kind, which have been met with throughout the whole animal kingdom and have been recorded in all parts of the world. These are arranged systematically under nearly nine hundred numbered headings, and are in many cases well illustrated by characteristic figures. The character and morphological relations of these variations are often very fully discussed with great knowledge and acuteness, and some original views are set forth which are of interest both to morphologists and physiologists. So far as this part of the work is concerned the present writer would feel himself quite incompetent to criticise it, but would welcome it as presenting in a convenient form a great body of interesting and little-known facts. But the book goes far beyond this. The first words of the preface tell us that "This book is offered as a contribution to the study of the problem of Species;" and in a lengthy introductory and shorter concluding chapters this problem is discussed in some detail, with the view of discrediting the views held by most Darwinians; while a new theory, founded upon the facts given in the body of the work, is set forth as being a more probable one.
It is therefore necessary to give some account of the nature of the facts themselves, as well as of the particular theories they are held to support.
Darwin distinguished two classes of variations, which he termed "individual differences" and "sports." The former are small but exceedingly numerous, the latter large but comparatively rare, and these last are the "discontinuous variations" of Mr. Bateson to which reference has been already made. Darwin, while always believing that individual differences played the most important part in the origin of species, did not altogether exclude sports or discontinuous variations, but he soon became convinced that these latter were quite unimportant, and that they rarely, if ever, served to originate new species; and this view is held by most of his followers. Mr. Bateson, however, seems to believe that the exact contrary is the fact, and that sports or discontinuous variations are the all-important, if not the exclusive, means by which the organic world has been modified. Such a complete change of base as to the method of organic evolution deserves, therefore, to be considered in some detail.
The difficulty which seems to have struck Mr. Bateson most, and which he declares to be of "immense significance," is, that while specific forms of life constitute a discontinuous series, the diverse environments on which these primarily depend shade into each other insensibly, and form a continuous series (p. 5). Further on, this objection is again urged in stronger language: "We have seen that the differences between Species are Specific, and are differences of kind, forming a discontinuous Series, while the diversities of environment to which they are subject are on the whole differences of degree, and form a continuous Series; it is therefore hard to see how the environmental differences can thus be in any sense the directing cause of Specific differences, which by the Theory of Natural Selection they should be" (p. 16). Again, at p. 69, he urges that the essential character of species is that they constitute a discontinuous series, and he asks—"Is it not then possible that the Discontinuity of Species
VOL. I. A A
may be a consequence and expression of the Discontinuity of Variation?" He then states, that on the received hypothesis, "Variation is continuous, and the Discontinuity of Species results from the operation of Selection." This, however, is not quite a correct statement of the received hypothesis if "discontinuous" is used in Mr. Bateson's sense, as including every change of colour which is not by minute gradation, and every change in number of repetitive parts—as of vertebræ, or of the joints of an antenna, or the rings of a worm—which is not by a gradation of the part from a minute rudiment. Such changes of colour or in the number of parts are admitted by all Darwinians as, in many cases, constituting a part of that individual variation on which modification of species depends. It is, however, on the supposed rejection of this class of variations by Darwinians that he bases what he terms "an almost fatal objection" to their theory.
Returning, however, to the supposed overwhelming importance of discontinuous variation, we pass on to the last chapter of the book, headed "Concluding Reflexions," and we read: "The first object of this work is not to set forth in the present a doctrine, or to advertise a solution of the problem of Species," and then follows immediately a further discussion of this very theory of discontinuity, which is set forth as a doctrine, and as a help to the solution of that problem. We are told that the difficulties of the accepted view "have oppressed all who have thought upon these matters for themselves, and they have caused some anxiety even to the faithful"; it is urged that "the Discontinuity of which Species is an expression has its origin, not in the environment nor in any phenomenon of Adaptation, but in the intrinsic nature of organisms themselves, manifested in the original Discontinuity of Variation"; that, "the existence of sudden and discontinuous Variation, that is to say, of new forms having from their first beginning more or less of the kind of perfection that we associate with normality, is a fact that disposes, once and for all, of the attempt to interpret all perfection and definiteness of form as the work of Selection." And then comes the positive statement—"The existence of Discontinuity in Variation
is therefore a final proof that the accepted hypothesis is inadequate" (page 568), and after several more pages of illustration and argument, the final conclusion is reached that—"it is quite certain that the distinctness and Discontinuity of many characters is in some unknown way a part of their nature, and is not directly dependent upon Natural Selection at all."
Before going further, it will be well to make a few observations on these very definite and positive conclusions at which Mr. Bateson has arrived; and it must be remembered that this volume deals only with one portion of the subject even of discontinuous variation, which is itself, if we exclude monstrosities, only a small fragment of the whole subject of variation. The impression that will be produced on those who have given special attention to the relations of living organisms to each other and to their inorganic environment, will be that of an academic discussion, dealing to a large extent with words rather than with the actual facts of nature. The author's main point, that species form a discontinuous series, and that specific differences cannot therefore have been produced by any action of the environment, because that environment is continuous—an argument which, as we have seen, he dwells upon and reiterates with emphasis and persistency—rests wholly upon the obvious fallacies that in each single locality the environment of every species found there is the same, and that all change of environment, whether in space or time, is continuous. To take this latter point first, nothing can be more abrupt than the change often due to diversity of soil, a sharp line dividing a pine or heather-clad moor from calcareous hills; or to differences of level, as from a marshy plain to dry uplands; or, for aquatic animals, from the open sea to an estuary, or from a non-tidal stream to an isolated pond. And when, in the course of geological time, an island is separated from a continent, or volcanic outbursts build up oceanic islands, the immigrants which reach such islands undergo a change of environment, which is in a high degree discontinuous.
Even more important, perhaps, is the fact that everywhere, the environment as a whole is made up of an
A A 2
unlimited number of sub-environments, each of which alone, or nearly alone, affects a single species, as familiarly included in the term, "their conditions of existence." The mole and the hedgehog may live together in the same general environment, yet their actual environments are very different owing to their different kinds of food, habits, and enemies. The same thing applies to the rabbit and the hare, the rook and the crow, the ring-snake and the viper; and still more when we look at animals of greater diversity, as the otter and the badger, the dung-beetle and the cockchafer, and a hundred others that might be quoted. Now, though all these creatures may be found together in the same area, each of them has its own "environment," to which it must be adapted in order to maintain its existence. Many species, however, live, as it were, on the borders of two distinct environments, as when they obtain different kinds of food at different periods, being then exposed to different enemies and varied climatic effects. In such cases, it is easy to see that a small modification of structure might enable them advantageously to change their habits, and thus obtain what would be practically a different environment. This is well seen in those closely allied species which have somewhat different modes of life—as the meadow pipit (Anthus pratensis) and the tree pipit (Anthus arboreus)—the former having a long, nearly straight claw to the hind toe, a more slender bill, and a rather greener tinge of colouring, all modifications suited to its different habits and distinct physical surroundings. Here we have an example in nature of how environments, even when continuous as a whole, may become quite discontinuous in relation to two species differing in very slight characters. Darwin dwelt much upon this phenomenon, of new species being formed when any body of individuals seized upon vacant places in the economy of nature, and by means of comparatively slight variations became adapted to it. It is what we see everywhere in the world around us.
Discontinuous Variations.
It thus appears that what is evidently supposed to be a very powerful argument leading to the conclusion that discontinuous variations as a class are those which are of vital, if not exclusive importance in the production of new species, entirely breaks down when confronted with the facts of nature. It does not, however, follow that, because an unsound a priori argument has been used to call attention to these variations, and because they have been set before the world in a way to suggest that their importance in relation to the origin of species is a new discovery calculated to revolutionise the study of this branch of biology, they are, therefore, of no value in this connection. We will, therefore, now proceed to consider them on their own merits as possible factors in the process of organic evolution. For this purpose we must briefly indicate the nature of the variations so laboriously recorded in this volume.
These consist of what are termed meristic variations, that is, variations in the number or position of parts which occur in series, whether linear, bilateral, or radial. Such are the variation in the number of segments of annulosa and arthropoda, such as worms, leeches, centipedes, &c.; in the antennæ and legs of insects; in the vertebræ, ribs, teeth, nipples, limbs, and toes of vertebrates; in the rays of starfish, encrinites, and allied animals. The ocelli and other symmetrical markings on the wings of butterflies are also recorded, as well as numerous malformations when these affect serial or symmetrical organs.
On carefully looking through the cases of variation in this volume we are struck with the large proportion of them which exhibit more or less deformation or want of symmetry, culminating in the various kinds of monstrosity. In Chapter III., on the variations of vertebræ and ribs, we find vertebræ imperfectly divided in snakes and frogs. Numerous cases of abnormalities in human vertebræ are given, usually exhibiting asymmetry or deformation, and similar variations are found in the anthropoid apes, but
here there is apparently more of regularity and symmetry. The greatest amount of this kind of variation occurs in the sloths, as might be expected when we consider that they are the most abnormal of mammals as regards the cervical vertebræ. In Chapter VIII. numerous cases of supernumerary mammæ are recorded, almost all of which are unsymmetrical. The variations in the number or form of the horns in sheep, goats, and deer recorded in Chapter XI. show them to be usually more or less irregular.
Nearly a hundred pages are devoted to the digits (fingers and toes) of mammals and birds, about one hundred and forty cases of variation being recorded. Almost the whole of these present, more or less, want of symmetry, while a large proportion, as the double-handed and double-footed children, and the six or seven-toed cats, can only be classed as monstrosities.
In succeeding chapters the variations in the antennæ and leg-joints of insects; in the radial parts of medusæ and encrinites; in the medial structures of fish, insects, molluscs, &c., which become sometimes double; in the eyes and colouration of flatfish; in duplicate or branching legs of insects and crustaceans; in extra limbs of batrachia; and, lastly, double monsters, are all discussed at great length, and are illustrated by a number of very interesting wood-cuts. But almost the whole of these can only be classed as malformations or monstrosities which are entirely without any direct bearing on the problem of the "origin of species."
Monstrosities have no Bearing on the Origin of Species.
Nothing can better show the small value of the book from this, which is the author's own, point of view, than the large amount of space devoted to the various monstrosities of the hands and feet of man and of some of the mammalia. Not only throughout all mammals, but also in the case of birds, reptiles, and amphibia, five is the maximum number of the toes or fingers. These may vary in size or in proportions; they may be reduced in number by coalescence, or by the loss of the lateral digits; they
may be strangely modified in form and function, as in the flappers of the whale, or in the wing of the bat, yet never once in the whole long series of land-vertebrates do they normally exceed five in number. Yet we have six, seven, or eight-fingered, double-handed, or double-footed children; similar malformations in monkeys; six and seven-toed cats; four, five, or six-toed pigs; double-footed birds, and other monstrosities, described at great length, and all their peculiarities discussed in the most minute detail and from various points of view, in a work presented to us as "a contribution to the study of the problem of species." Many of these malformations have been observed among animals in a state of nature, and in fact, Mr. Bateson believes that they occur as frequently among wild as among domesticated animals. Considering how rarely the former cases can be observed, they must be everywhere occurring; yet in no one single instance do they seem to have established themselves as a race or local variety on however small a scale. Yet we know that in the case of the six-toed cats, and probably in other cases, they are easily transmissible; and we must, therefore, conclude that all these irregularities and monstrosities are in a high degree disadvantageous, since when subject to free competition with the normal form in a state of nature they never survive, even for a few generations.
Variations in Number of Serial Parts rarely a Factor in Origin of Species.
As the volume we are discussing is entirely devoted to variations in the number or position of the serial parts of organisms in relation to the origin of species, it becomes necessary to lay some stress upon the very familiar, but apparently overlooked fact, that, among all the higher types of life at all events, the most stable of all characters, and the most permanent during long periods of evolution, and throughout changes which have led to the production of a marvellous variety and abundance of specific forms, are these very characters of the number and relative positions of serial organs; whence it follows that variations
of this kind can only have led to specific changes at enormously long intervals, and that, as a general rule, they can have had nothing whatever to do with the origin of an overwhelming majority of living species.
First, we have the four limbs of vertebrates, which among all the marvellous variety of form and function, on land, in the water, or in the air, is never exceeded, and appears to have been fixed at a very early stage of the development of the vertebrate type. Equally fixed, and extending through a still vaster range of modifications of specific forms, are the six legs and four wings of true insects, which, as in vertebrates, may be reduced but never increased in number. Still more extraordinary, because less obviously connected with the main structure and functions of the organism, is the limitation and permanence in the number of the subdivisions of limbs and other appendages. There is no obvious reason why in land-vertebrates the divisions of the hand and foot should never exceed five, yet, not only is this number the maximum, but it may be considered the normal number of which all others are reductions, since it still prevails largely in the marsupials, rodents, carnivores, primates, and lizards; and the five-toed land-vertebrates (excluding birds) are probably far more numerous than those with a lesser number.
In birds there are only four toes as a maximum, and comparatively few have a smaller number. But we have here a peculiarity in the numbers of the toe-joints which does not occur in any other vertebrates. These form a series in arithmetical progression, the hind toe having two, and the others three, four, and five joints in regular order; and this rule is very nearly universal, the only exceptions being in some of the swifts and goatsuckers, whose habits render the feet of comparatively little importance, while their general organisation is of a somewhat low type.
Coming to insects, we again find the legs consisting of a limited number of parts, and, strangely enough, this number is again five—the coxa, trochanter, femur, tibia, and tarsus. The tarsus, however, is subdivided into small movable joints, and these, too, are five as a maximum, but
in certain groups are reduced to four, three, or two. The five-jointed tarsus is, however, the most prevalent; and in the enormous order of coleoptera or beetles, comprising at least one hundred thousand described species, fully half belong to families which have the tarsi five-jointed. Even the antennæ, although they vary greatly in the number of joints, yet in numerous large groups comprising many thousands of species, they have the number of joints constant. Another indication of the tendency of serial parts to become fixed in number, is the typical limitation of the cervical or neck vertebræ of mammalia to seven joints. This number is wonderfully constant, being the same in the long necks of the giraffe and camel, and the very short necks of the hippopotamus, porpoise, and mole, the only exceptions in the whole class being some of the sloths, which have from six to ten, often varying in the same species, and the manatee, which has six.
Now, if we consider the enormous extent of these fixed numerical relations of important parts of the organism in the higher vertebrates and in insects, both as regards the number of living species affected—perhaps ninety-nine per cent. of the whole—and as regards their range in time, throughout the whole of the tertiary and secondary, and even a considerable portion of the palæozoic periods—and if we take account of the vast number of extinct species, genera, and families needful to complete the various lines of descent from the earliest known forms, presenting the same numerical relations to those now living—we shall be able to form some conception, however inadequate, of the overwhelming frequency and importance of variations in the size, form, proportions, and structure of the various parts and organs of the higher animals, as compared with variations in their number. No doubt, in the earlier stages of organic development, numerical variations were more frequent and more important, as they are now among the lower forms of life, but at a very early period in geological history, the main numerical relations of the essential parts of the higher organisms became more or less fixed and stable, and have in many cases remained
unchanged through a large proportion of the period comprised in the geological record. The four limbs of vertebrates were already established in the fishes of the Devonian period, as were the four wings and six legs of true insects in the cockroaches and archaic orthoptera of the Carboniferous; and almost all subsequent changes have resulted from modifications of these early types. The earliest mammals of which we have sufficient knowledge have the typical five-toed feet, and the earliest birds appear to have had the same progressive series of toe-joints as now prevails.
We are thus irresistibly led to the conclusion that, among all the possible forms of variation now occurring, those affecting the number of important serial parts among higher organisms, are those which have the least possible relation to whatever modification of species may now be going on around us, or which has been going on during a large portion of geological time. Yet it is to variations of this nature, a large proportion of which are mere malformations or monstrosities, that the bulky and learned volume we are discussing has been devoted. The author of this book puts forward these malformations and irregularities, mixed up with a proportion of normal variations, under the misleading name of "Discontinuous Variations," as if they were something new, and had been ignorantly overlooked by Darwin and his followers; and he loses no opportunity of telling us how important he thinks they are, what difficulties they enable us to overcome, and how they are the beginnings of the establishment of a sure base for the attack on the problems of evolution. In so doing he has entirely failed to grasp the essential features which characterise at least 99 per cent. of existing species, which are, slight differences from their allies in size, form, proportions, or colour of the various parts or organs, with corresponding differences of function and habits, combined with a wonderful amount of stability in the numerical relations of serial parts, extending sometimes only to genera, but more usually to families, tribes, orders, or even to whole classes of the
higher animals. It is differences of the former kind that do actually characterise the great majority of species;1 they affect those organs which vary most frequently and most conspicuously in the individuals of every fresh generation; and they constitute that "individual variation" on which Darwin always relied as the essential foundation of natural selection, and which his followers have shown to be far more abundant and of far greater amount than he was aware of; and, lastly, they afford amply sufficient material for the continuous production of new forms. Rarely in the history of scientific progress has so large a claim been made, and been presented to the world with so much confidence in its being an epoch-making discovery, as Mr. Bateson's idea of discontinuous variation corresponding to and explaining the discontinuity of species; yet more rarely has the alleged discovery been supported by facts which, though interesting in themselves, are for the most part quite outside the general conditions of the complex problem to be solved, and are therefore entirely worthless as an aid to its solution.
Before leaving this part of the subject we may note the extension of definite numerical relations to plants as well as to animals. In dicotyledons we have a typical five-petalled flower or a corolla with five divisions, a character which prevails in irregular as well as in regular flowers, and often when the stamens are not a multiple of five, as in mallows, bignonias, and many others. Some form of five-parted flower prevails throughout many extensive natural orders, and comprises probably a considerable majority of all dicotyledonous plants. A three or six-parted flower is almost equally a characteristic of monocotyledons, prevailing even among the highly specialised and fantastically formed Orchideæ and Irideæ, thus again demonstrating how large a portion of the specific modifications of organisms are independent of variations in number, but depend wholly upon variations
1 Mr. Bateson, however, makes the extraordinary statement that "it is especially by differences of number and by qualitative differences that species are commonly distinguished" (p. 573). Species-makers know too well that, among the higher animals at all events, it is not so!
in the size, form, colour, and structure of the various parts and organs.
Other matters of importance in Mr. Bateson's work, together with some theories recently advanced by Mr. Francis Galton, will now be discussed.
Some Minor Objections to the Darwinian Theory.
We will now deal with some of the minor objections to the views of most Darwinians which are to be found in Mr. Bateson's lengthy Introduction; after which the validity of Mr. Francis Galton's doctrine, as to positions of organic stability (also held by Mr. Bateson), will be considered. And first, we note that he uses the usual misleading terms, "minute," "minimal," "imperceptible," and "insensible" (p. 15) as applied to the individual variations on which Darwin relied, although he has himself given figures of beetles and earwigs showing that such variations are enormous—greater, indeed, than in the illustrative cases I have given in my Darwinism.1
A strong attack is made on the theory of the utility of specific characters. It is admitted that an enormous amount of evidence has been collected, and that "the functions of many problematical organs have been conjectured, in some cases perhaps rightly"—yet he adds, "whole groups of common phenomena are still almost untouched even by conjecture." He tells us that "many suggestions have been made as to the benefits which edible moths may derive from their protective colouration; and as to the reasons why unpalatable butterflies in general are brightly coloured" (p. 11)—but neither here nor elsewhere is any hint given that more than "suggestions" have been advanced. Considering that this is the one branch of the subject in which natural selection has been shown to be an actual working reality in nature by the experiments of Jenner Weir, Butler, Stainton, and Belt, the observations of Bates and Fritz Müller, and especially by the experimental investigations of Professor
1 See Proceedings of the Zoological Society, 1892 (p. 59—23), in a paper by W. Bateson and H. H. Brindley.
Poulton, it was hardly fair to pass the subject by as if nothing had been done but pure conjecture. He also ignores the continuous advance that is being made in determining the utilities of the innumerable modifications in the forms and arrangement of the leaves and other of the non-floral appendages of plants by Kerner, Lubbock, and many other observers; as well as the light thrown on colour and marking as specific characters in the higher animals by the consideration of the value of distinctiveness for purposes of recognition, a character of life-preserving value in the case of many animals, and in all of great importance to reproduction, and an essential factor in the differentiation of species. It is, therefore, not correct to say, "But as to the particular benefit which one dull moth enjoys as the result of his own particular pattern of dulness as compared with the closely similar pattern of the next species, no suggestion is made." The suggestion has been made (Darwinism, p. 226), and has been accepted as at all events a good working hypothesis by many naturalists. On this question of the utility of characters which are constant characteristics of the species, but whose utility is not apparent to the casual observer, Mr. Bateson uses very strong language. Referring to the case of two ladybirds, the small Coccinella decempunctata being exceedingly variable, both in colour and spotting, the larger, C. septempunctata, very constant, he says, "To be asked to believe that the colour of C. septempunctata is constant because it matters to the species, and that the colour of C. decempunctata is variable because it does not matter, is to be asked to abrogate reason" (p. 572). I fear that I myself must be in this sad case, for though I have not been asked to believe this unreasonable thing, yet I do believe it! Of course I may be wrong and Mr. Bateson right, but how is it that he is so absolutely sure that he is right?
Before proceeding further we may briefly notice that Mr. Bateson seems to imply that the "meristic," or numerical variations, to which he has devoted his volume, are altogether ignored by Darwinians in their adoption of "individual variations" as opposed to "sports" for the
main materials on which natural selection works. But this is altogether erroneous. No doubt they would reject nine-tenths of Mr. Bateson's cases as being simply monstrosities, which neither have nor could ever have had any part in the production of new species; but they always recognise that genera, and even species are sometimes characterised by a difference in the number or arrangement of serial parts—as of vertebræ, ribs, teeth, or markings, and that variations of this kind do sometimes, though in comparatively few cases, form the raw material on which natural selection works. As development seems almost always to have proceeded by reduction from large and indefinite numbers of serial parts to the minimum number compatible with the maximum of utility, any increase in number occurring now, may be, as is usually considered, a form of reversion, though Mr. Bateson denies that there is any such thing in nature. This diminution in number may have occurred either by a gradual diminution in size and ultimate disappearance, as when limbs of the higher animals have been lost, in whales, the apteryx, snakes, &c.; or it may sometimes have been abrupt, which means that the rudiment of the part ceased to develop at an early embryonic stage. Either mode is quite in harmony with the views of Darwinians, and not very much seems to be gained by terming the former "continuous" and the latter "discontinuous," especially when this last term is held to include almost every kind of monstrosity.
Organic Stability as a Factor in Evolution.
We have now to consider an equally important, though, as I think, an equally unsubstantial novelty—the view that there are "definite positions of organic stability," which alone are sufficient to mould races "without any help whatever" from natural selection. This view appears to have originated with Mr. Francis Galton, and was first stated in his work on "Natural Inheritance," and again in his Royal Society paper on "Thumb and Finger Marks." The same view is adopted by Mr. Bateson; and in an article on "Discontinuity in Evolu-
tion" in Mind (vol. iii., pp. 362, 372) Mr. Galton approves of Mr. Bateson's work, and restates his latest views on the subject, and these I now propose to consider.
Although Mr. Galton begins by the consideration of races only, that is, of well-marked forms below the value of species, yet later on he applies his theory to the evolution of species and of all higher groups. Speaking of discontinuous or, as he terms it, transilient variation, he says: "A leap has taken place into a new position of stability. I am unable to conceive the possibility of evolutionary progress except by transiliences, for, if they were mere divergences, each subsequent generation would tend to regress backwards towards the typical centre, and the advance which has been made would be temporary and could not be maintained" (l. c., p. 368). Mr. Galton has before implicitly admitted that there is such a thing as natural selection, yet in this passage he reasons as if it had no existence, and that regression to mediocrity would occur just the same with it as without it. For the essence of natural selection is that it preserves and thus increases favourable variations by destroying the unfavourable; but this statement by no means expresses the real power of selection, which may be better illustrated by saying that it destroys about 99 per cent. of the bad and less beneficial variations, and preserves about the 1 per cent. of those which are extremely favourable. With such an amount of selection how can there be any possible "regression backwards towards the typical centre" whenever any change in the environment demands an advance in some special direction beyond it as the only means of preserving the race from extinction? Well did Darwin say that unless the universal struggle for existence "be thoroughly ingrained in the mind, the whole economy of nature, with every fact on distribution, rarity, abundance, extinction, and variation will be dimly seen or quite misunderstood." Almost all the misconceptions of popular writers against Darwinism arise from the want of this constantly present recollection; and as capable and instructed writers, such as Mr. Galton and Mr. Bateson, as well as capable but uninstructed writers,
such as Lord Salisbury, alike suffer in this respect, it is needful to again state obvious facts which may serve to drive home the overwhelming importance of this factor in evolution.
Let us suppose an animal which lives ten years and produces ten young (five pairs) each year, a moderate allowance even for many mammals and birds. A little simple arithmetic will show that if none died for five years there would be 6,480 pairs in place of the one pair, or, 6,480 millions in place of one million, as the case might be. But it is evident that such an average rate of increase for all animals could not go on for even one or two years, as no country could supply them with food. We will suppose, then, that only one pair, instead of five, survive each year to breed the next year; but if this goes on for the ten years of the life of the first pair we shall still have 512 pairs instead of each pair, a number which is equally impossible. Let us, then, suppose that only one-fiftieth part of those born survive, that is, that only one individual lives to breed out of five successive broods of ten each; even then, at the end of ten years, we shall have a population two and a half times as great as at first, or, more exactly, if we began with a million individuals, then in ten years we should have 2,593,743. This is probably something like what happens. Forty-nine fiftieths of those born never live to breed, yet the population increases steadily so long as conditions are moderately favourable, the surplus being got rid of at uncertain intervals by recurrent unfavourable conditions, so as to keep the number of individuals on the average about stationary. Looking at it in another way, we find that, beginning with 100 individuals whose offsprings each year amount to 500, of which only 10 survive to breed, then during ten years about 8,000 will have been born, making with the original hundred, 8,100, out of which only the 100 fittest, or nearly the fittest, will survive, to be again weeded out every successive ten years, or thereabouts. Without making some numerical estimate of this kind it is impossible to realise the severity of the struggle continually going on in nature and the resultant
elimination of the unfit. With the above figures (which would have to be enormously greater with many species) we see that for every 80 born only 1 on the average survives to breed. With such an amount of selection it is evident that whenever it happened that the mean point, or "typical centre" of the curve of variation, ceased to be the most advantageous point in relation to the whole conditions of existence, then a new typical centre would rapidly be produced by the elimination of all which diverged from it to any injurious extent. There could not possibly be any permanent regression from the new typical centre unless the inevitable survival of the fittest in a rapidly increasing population can be got rid of.
We are now in a position to discuss Mr. Galton's theory, that there are certain variations which possess "organic stability," and that these are the real factors of evolution "without any help whatever from the process of selection." And first, what is the exact character of these stable variations, which form races and ultimately new species by their own inherent force of stability? Is the stability in relation to the actual conditions of the environment or altogether independent of those conditions? If the former, how did it come to be in harmony with them? If this harmonious relation depends upon a mere chance coincidence, we have to consider the comparative rarity of these large or discontinuous variations, and that only a small proportion of them have the alleged character of "stability." Moreover this class of variations is generally a variation in a single part or organ, and Herbert Spencer (as well as many other writers) has argued forcibly that modifications of single characters would in all cases be useless unless accompanied by the correlative modifications of a number of other characters. I have myself shown that in the case of ordinary variations this is no difficulty, because all characters are varying more or less in every generation, and thus the needful harmonious relation between the different organs or parts can be easily maintained; but in the case of these large exceptional variations the difficulty is an overwhelming one. And we must always remember that these alleged "stable" vari-
VOL. I. B B
ations, from the first moment of their appearance, cannot possibly escape from the action of natural selection. As, roughly speakly, only about one per cent. of each generation survives to breed, this new form, however stable in itself, cannot become part of that one per cent. unless the particular variation which characterises it is either beneficial or entirely harmless. But in the latter case it is difficult to see what constitutes its superiority over the rest of the species which, year by year, by means of this intensely severe process of elimination, is kept in harmony with the environment. If the stability consists in greater health, vigour, reproductive power, or intelligence, all these qualities have already been developed to the fullest needful extent, and these superior individuals will be selected in the usual way. But if—as a second alternative—these alleged stable variations are supposed to have some kind of inherent stability independent of the environment, then, stability notwithstanding, they would inevitably soon succumb under the terrible eliminating power which year by year leaves only about one per cent. of the most fit to survive. There is really no escape from this dilemma: If your new variety is among the one or two per cent. of the most fit, then it does not need this purely imaginary quality of "organic stability" in order to survive; if it is not among this small body of the most fit—that is, of the best adapted to the whole conditions of existence of the species or race—then, any other quality notwithstanding, it will certainly not survive. The term "organic stability" has really no meaning except that of harmonious adaptation to the environment as tested and maintained by natural selection. To talk about new races or species being produced "without any help whatever from natural selection" can, under the actual conditions of the terrestrial universe, only mean that there is an inherent developmental power which modifies organisms in definite ways, and in more perfect harmony with the environment than has been, or can be, brought about by natural selection, thus keeping these modified organisms always ahead of the rest. They may thus be said to be independent of survival of the fittest, because they and
their offspring always are the most fit, and therefore always survive! On this theory evolution goes on by the production of new races complete and ready formed, and in perfect harmony with the environment whenever that environment undergoes a change. But no evidence is offered for such an extraordinary developmental power being always at work and always able to produce adaptation to an ever changing environment. Such a power would be hardly different from the old special creation, or than the pre-ordained harmony of the philosophers; and it would, moreover, have rendered unnecessary and unintelligible that rapid multiplication, and consequent enormous expenditure of life, which now prevails. It would equally render unnecessary that wonderful property of individual variability, whose only use would then be to enable man to improve his domestic animals and cultivated plants. We should thus have two rival systems at work, and we might almost imagine Mr. Sullivan's cosmic spirits—William and James—to be realities, and that each had been experimenting in organic development on our earth in order to see whose scheme was the most satisfactory.1
Finger-marks as Illustrating Organic Stability.
As evidence of the actual existence of this hypothetica "organic stability," Mr. Galton adduces the patterns in thumb and finger-marks, which he has so carefully studied. In his Royal Society paper on this subject, he tells us that these marks fall into definite groups and can be systematically classified, and he actually describes and figures twenty-five distinctive patterns arranged under three very distinct classes. He then urges that these fundamentally distinct classes are strikingly analogous to genera in biology, and as the patterns are so insignificant in every way that they can in themselves be neither useful nor ornamental, and can therefore never have been the subjects of selection, they prove, he thinks, "that natural selection has no monopoly of influence in forming
1 See Strand Magazine, vol. iv.
B B 2
genera, but that it could be wholly dispensed with, the internal conditions acting by themselves being amply sufficient to form them." And it is from the case of these finger-marks that he considers the reality of positions of organic stability has been proved, and that they are "competent to mould races without any help whatever from the process of selection."
At first sight this may appear to be sound reasoning, and to be fatal to some of the claims of the Darwinians, but further examination will show that it is a pure fallacy arising from the vague use of terms, and from comparing quite different things as if they were of the same nature. The fallacy depends on applying the terms of classification in systematic biology to groups of single objects which have no real relation with the genera and species of the naturalist. The essential character of a species in biology is, that it is a group of living organisms, separated from all other such groups by a set of distinctive characters, having relations to the environment not identical with those of any other group of organisms, and having the power of continuously reproducing its like. Genera are merely assemblages of a number of these species which have a closer resemblance to each other in certain important and often prominent characters than they have to any other groups of species. It will be more intelligible and more instructive if we confine ourselves to species as the unit of comparison with Mr. Galton's groups of stable finger-patterns, in order to show the fundamental differences between them. And first we see that Mr. Galton classifies the marks themselves, not the individuals who possess the marks. He tells us that the very same general varieties in these marks are found in English, Hindoos, and Negroes, and, presumably in all other races; and, further, that he has "failed to observe any correlation between the patterns and any single personal quality, whether mental or physical." All this is entirely different from either specific or generic characters, whose essential feature is that they are found in every normal individual of the genus or species, and are always correlated with other characters. In his first paper on
this subject (in 1890), Mr. Galton said he had reason to believe that the patterns are to some extent hereditary, but that he had no evidence of it; while in his paper in Mind four years later, he could still only say that "they are to be looked upon" as having "a slight tendency towards transmission by inheritance." But the very essence of specific and generic characters is, that they are strictly transmitted by inheritance. Yet again, whatever difference of opinion there may be as to the utility of all the characters which distinguish species, every one will admit that many are useful, and especially that the general assemblage of characters that fit each species for a somewhat different mode of life from its nearest allies, must certainly be useful. But the very essence of Mr. Galton's argument as to these finger-prints is, that they are not and cannot be in any way directly useful. How, then, can the manner in which these patterns may be grouped, furnish us with any argument whatever as regards such totally diverse things as generic or specific characters—and still less as regards genera and species themselves?
The fact is, no doubt, that these patterns are the direct result of the laws of growth of the tissues of the skin. The limited number and definite character of these patterns are probably the mechanical incidental results of these laws, under the ever-varying conditions of development in each individual. A good analogy would be found in snow-crystals, of which about a thousand varieties have been recorded, which may, however, all be grouped under five classes, while each snow-fall usually produces crystals of one class. Here we have the fixed and definite laws of the crystallisation of water, so modified by conditions of moisture, temperature, motion, and perhaps electric state of the atmosphere, as to lead to this wonderful variety of the product, yet always subject to the law of crystalline symmetry and to systematic grouping under definite classes; just as in these finger-prints we have a more limited variety of forms, which also can be grouped under a few classes. But neither the one nor the other has any real bearing upon the problem of the nature and
origin of the genera and species of living organisms. A study of the distribution of the stars over the surface of the heavens, or of the interlacing ripple-marks upon the sea-beach, would no doubt show that these objects might also be the subject of classification; and from the point of view of elucidating the origin of species, they would be about as useful, or as worthless, as the study of finger-marks.
Of course, there are many varieties or races, both among animals and plants, which continually reappear, and which in some cases are known to reproduce their like, and these undoubtedly have an appearance of stability. Such are the light and dark-coloured varieties in many insects and in some mammals and birds; the hairy or smooth varieties of plants; specially banded or coloured land-shells, and many others. Whenever any of these variations are not injurious under the actual conditions of existence of the species, they may persist in considerable numbers, and thus appear to be stable. But others which are comparatively rare may be just as stable organically, as shown by the case of white mice, pigeons, &c., which increase to any extent under domestication. In a wild state they never do so, and the obvious reason is, that either the conspicuous colour, or something correlated with it, is injurious. In flowers white varieties are frequent, and they occur in all degrees of abundance or rarity; and this indicates, in all probability, various degrees of hurtfulness. If in any case the white colour were not at all injurious as compared with that of the type, it would almost always, by the operation of Delbœuf's Law, tend to increase to nearly an equality with the parent form; and as this equality so rarely occurs, we must conclude that, in most cases, the variety (of whatever kind) is to some extent injurious.1 From the usually limited num-
1 For a statement and popular demonstration of Delbœuf's Law, see Habit and Intelligence, by J. J. Murphy, 2nd Ed., p. 241. Briefly, the law is, that, if a species produces a variety, in however small a proportion annually, and if the variety produces its like in the same proportion as does the species, and if it is neither beneficial nor hurtful to the species, then the variety will increase, rapidly at first and more slowly afterwards, till it approaches to equality in numbers with the species. From this law it follows that, as varieties are usually very much less numerous than the species, this must be due to one of the following causes: either (1) the variety has but recently originated, and has not had time to increase; or (2) the variety has ceased to be produced by the species; or (3) it does not reproduce its like so completely as does the species; or (4) it is disadvantageous to the species. The first two suppositions are improbable, and can only account for a very small proportion of the varieties which are greatly inferior in numbers to the species; the other two are antagonistic to any special "organic stability," which must, therefore in the great majority of cases be rejected as being both unproven and also opposed to the facts.
ber of individuals presenting these discontinuous variations, we may therefore draw an important conclusion which has hitherto been overlooked. It is, that not only do such variations afford no support to the theory of a special "organic stability" capable of producing races, species, and even genera, without any aid from natural selection, but they furnish a strong, if not conclusive, argument against it, since any which did possess such exceptional stability, and were in no degree injurious, would long since have become equal in numbers to the type of the species.
Laws of Growth, Importance of.
A few words are here necessary as to the very common misconception that extreme Darwinians do not recognise the importance of the organism itself and of its laws of growth and development, in the process of evolution. For myself, I may say that no one can be more profoundly impressed by the vast range, by the complexity, by the mystery, by the marvellous power of the laws and properties of organised matter, which constitute the very foundation of all life, and which alone render possible its countless manifestations in the animal and vegetable worlds; while those who have read Weissmann's account of the complex processes of development of sperm and germ cells, in his volume on The Germ Plasm, must feel sure that he, at all events, can have no inadequate conception of their importance.
What Darwinians deny is—as I understand the question—that these laws themselves serve to keep the completed organism in close adaptation to the fluctuating
environment, instead of merely furnishing the material which is required for that adaptation. In our view, the fundamental laws of growth and development, through the agency of rapid multiplication and constant variability, provide the material on which natural selection acts, and by means of which it is enabled to keep up the adaptation to the environment (which alone renders continuous life and reproduction possible) during the constant, though slow changes, whether inorganic or organic, by which, in the course of ages, the effective environment of each species becomes more or less profoundly modified. Thus, and thus alone, we believe, are new species produced in strict adaptation to the new environment. So far as rendering possible and actually leading to growth, reproduction, and variation, the fundamental laws are supreme. In securing the development of new forms in adaptation to the new environment, natural selection is supreme. Hence arises the real distinction—though we may not always be able to distinguish them—between specific and non-specific or developmental characters. The former are those definite, though slight modifications, through which each new species actually became adapted to its changed environment. They are. therefore, in their very nature, useful. The latter are due to the laws which determine the growth and development of the organism, and therefore they rarely coincide exactly with the limits of a species. The more important of these latter characters are common to much larger groups, as families, orders, or classes, while others, depending partly on complex and fluctuating influences, are variable even within the limits of a species. Of this last kind are the finger-prints, which, like many other minute details of form or structure, vary from individual to individual.
I have now, I think, shown that the two most recent efforts to establish new "methods of organic evolution" as either complete or partial substitutes for natural selection—that is, for the survival of the fittest among the individual variations annually produced—have completely failed to establish themselves as having any relation to the
actual facts of nature. Mr. Bateson's discontinuous variations were long ago rejected by Darwin as having no important part in the formation of new species, while recent and ever-growing proofs of the generality and the magnitude of individual variability, render these larger and rarer kinds of variation of even less importance than in his time. Mr. Galton's theory of organic stability, which is essential to the success of discontinuous variations, has been shown to be founded upon a comparison of things of a totally dissimilar nature, and, further, to be absolutely unintelligible and powerless unless in strict subordination to natural selection.
The reason why two writers of such extensive knowledge and undoubted ability have so completely failed in dealing with the great problem of the modification of organic forms, has been clearly indicated during the course of this discussion. It has arisen from the fact that they have devoted themselves too exclusively to one set of factors, while overlooking others which are both more general and more fundamental. These are, the enormously rapid multiplication of all organisms during more favourable periods, and the consequent weeding out of all but the fittest in what must be on the whole stationary populations. And, acting in combination with this annual destruction of the less fit, is the periodical elimination under recurrent unfavourable conditions, of such a large proportion of each species as to leave only a small fraction—the very elect of the elect—to continue the race. It is only by keeping the tremendous severity of this inevitable and never-ceasing process of selection always present to our minds, and applying it in detail to each suggested new factor in the process of evolution, that we shall be able to determine what part such factors can take in the production of new species. It is because they have not done this, that the two authors, whose works have been here examined, have so completely failed to make any real advance towards a more complete solution of the problem of the Origin of Species than has been reached by Darwin and his successors.
CHAPTER XVIII
THE PROBLEM OF UTILITY: ARE SPECIFIC CHARACTERS ALWAYS OR GENERALLY USEFUL?1
THE above stated question is discussed at great length in the second part of the late Mr. Romanes' work on Darwin and After Darwin, fully half of the volume being devoted to it; and in the preface the author states his belief that his arguments are so conclusive that he has "broken to fragments" the doctrine of utility, and that he has "made a full end thereof." A careful perusal of the volume, and a full consideration of all the facts and arguments adduced therein, seem to me to leave the problem just where it was before; but the variety of the subjects discussed, the great mass of details referred to, and the ingenuity of some of the arguments in support of the author's view, lead me to think that I have not hitherto set forth the facts and arguments in favour of the utility-theory with sufficient completeness, while I am indebted to the lamented author for pointing out one or two weak points in my discussion of the question, and for a number of useful references to Darwin's statements on the points at issue, some of which I had overlooked.
Although Mr. Romanes' discussion of the question is so lengthy, the problem itself is in its essence a comparatively simple one, and is I believe capable of being solved by a reference to well-known facts and admitted principles. The reason why Mr. Romanes is able to support his views
1 This paper was read before the Linnean Society on June 18th, 1896, and printed in the Journal—Zoology, vol. xxv.
by so many quotations from Darwin's works, is due to the fact that Darwin was firmly convinced of the heredity of acquired characters, and especially of the influence of food and climate and the effects of use and disuse; and this belief must be borne in mind whenever he speaks of specific characters being due to other causes than natural selection. It must also be remembered that Darwin was not acquainted with the evidence we now possess as to the extreme frequency of variation everywhere in nature, its large amount, and its universality in every organ and every character that can be measured or otherwise estimated. Had he known what we now know on this subject, he would not so frequently have made the proviso—"if they vary, for without variation natural selection can do nothing," or have alluded to the possibility of variations of the same kind occurring "perhaps after a long interval of time." We now know that variations of almost every conceivable kind occur, in all the more abundant species, in every generation, and that the material for natural selection to work upon is never wanting. Accepting, then, these facts of variation, and always keeping in mind the severity of the struggle for existence, nine-tenths at least of the progeny of the higher animals perishing annually before reaching maturity, thus leading to a systematic and continual weeding out of the less fit—let us endeavour to realise the process of the formation of new species and the nature of the characters which distinguish allied species from each other.
The Principle of Utility.
In my article on "Mimicry and other Protective Resemblances among Animals," first published in 1867, I laid down the principle of utility, perhaps a little too absolutely, in the following passage:—"Perhaps no principle has ever been announced so fertile in results as that which Mr. Darwin so earnestly impresses upon us, and which is indeed a necessary deduction from the theory of Natural Selection, namely—that none of the definite facts of organic nature, no special organ, no characteristic form or
marking, no peculiarities of instinct or of habit, no relations between species or between groups of species, can exist but which must now be, or once have been, useful to the individuals or races which possess them." Professor Huxley, in his obituary notice of Darwin, expressed the same idea as follows:—"Every variety which is selected into a species is favoured and preserved in consequence of being, in some one or more respects, better adapted to its surroundings than its rivals. … For, as has been pointed out, it is a necessary consequence of the theory of Selection that every species must have some one or more structural or functional peculiarities, in virtue of the advantage conferred by which it has fought through the crowd of its competitors and achieved a certain duration. In this sense it is true that every species has been 'originated by selection.'" Now these characters, in virtue of which the variety has become a species, are in fact its "specific characters," and they alone will absolutely differentiate it from all other species. We need not trouble ourselves about the cases of doubtful species, in which the distinctive characters are either so minute or so unstable that we cannot invariably determine them. On the theory of evolution by natural selection there must be such cases. They are species in the making and not quite completed. But in the great majority of species definite characters do exist by which any single individual can be recognised and the species to which it belongs be determined; and the question is, whether or no the characters, or combination of characters, which thus differentiate it are now useful or were useful at the time of its origination.1 In order to answer this question, we must briefly summarise both the facts and the admitted principles or theories which bear upon it.
1 To this should be added—"or were correlated with some useful characters." I have referred to such correlations in my Natural Selection and Tropical Nature, pp. 172 and 175; and as to apparently useless characters being in some cases correlated with those which are useful, in my Darwinism, p. 140; but it is cumbersome to restate this part of the theory whenever it is alleged that all specific characters are useful.
Why Species are Stable
Every extensive area contains a number of large and dominant species which appear to be, and probably are for considerable periods, stable, both in average population and in the extent of the area they occupy. Taking any one of these species—say of bird or mammal—so long as the whole conditions of its environment remain unchanged or very little changed it will, theoretically, continue to maintain itself, as we know many species have maintained themselves during the whole period since the glacial epoch, and some very much longer. The species, however, is not absolutely homogeneous. It varies in every generation, not minutely or infinitesimally as was formerly supposed, but very considerably, the variations being easily seen and measured by any one who looks for them; and they extend, so far as we know, to every part of the organism, external and internal, since no part has yet been found to be invariable when a large number of individuals have been compared. The species is therefore composed of a fluctuating mass of variable units which yet maintain the same general average or characters, and this it can only do by a constant or intermittent weeding out of the extremes in every direction. Such a weeding out on a large scale takes place annually, because, although the annual increase by birth is very large, the population of adults remains approximately fixed. The species is maintained in harmony with its environment by the survival of the fittest.
Why Species become Modified.
But now let some important change occur, either in climate, in abundance of food, or by the irruption of some new and hitherto unknown enemies, a change which at first injuriously affects the species. It must, therefore, undergo some amount of modification, either structural or functional, in order to succeed under the new conditions; and the constant variations of every part (around its mean) furnish the materials for adapting the organism to these new conditions. If a new enemy is the danger to be guarded against
this adaptation may be effected in several ways. Swiftness in running or flying, habits of concealment, or seeking new kinds of food in places inacessible to the enemy, may each lead to the survival of those individuals which were sufficiently intelligent to adopt them, or sufficiently favoured by rapid variation in the desired direction. Survival of the fittest in these respects, going on year by year, might lead to the formation of two or more diverging races each able to maintain itself in the presence of the new enemy, while the former average type of the species rapidly became extinct. We should thus have two or three incipient new species; but they would not become well differentiated species till they had acquired certain definite and important characteristics. These are (1) some amount of infertility when crossed with the parent form or with each other; and (2) some distinct and conspicuous external characters by means of which the new varieties could readily distinguish their own kind even when at considerable distances or when partially concealed; or, in the case of flowering plants, be distinguished by the insects which fertilise them.
The greatest danger to a species under new and adverse conditions is, that it should not be able to adapt itself to them with sufficient rapidity. It is for this reason that, as Darwin concludes, new species arise, mainly, from those which have a large population, which occupy a wide area, and which present much variation—a combination rarely found except in the great continents. But this danger is evidently much increased if crossing with the parent form is not at first checked and soon afterwards completely prevented except as a quite exceptional occurrence. The means of preventing this intercrossing are, for animals, either infertility, external distinctions leading to the preferential mating of similar forms, or physical isolation. The latter I believe, with Darwin, to be of comparatively little importance and to have very rarely been the chief agent in modification. In the great majority of cases a new species must arise amidst the population of an existing species; and while its adaptation is progressing any intercrossing with the parent form will be injurious. I have endeavoured to show, and can still find no flaw in my reasoning, that
mutual infertility would be usually brought about by natural selection wherever the two forms were in contact, and also that the early occurrence of well-marked external differences would assist greatly in the rapidity of adaptation.1 This view will explain the curious fact of the well-marked differences of colour or form which almost invariably characterise allied species. These "recognition marks," as I have termed them, are of great use even to existing well-defined species, but they must have been of still greater use during the earlier stages of differentiation, when the very existence of the new form must have largely depended on them.
I may here remark that it is because these external differences of colour or marking are quite as constantly present in peculiar insular species as in those inhabiting a continent, that I do not believe in local isolation as of any real importance in species-formation. Insular species may have been produced in two ways. Either a portion of a declining species may have reached the island, where it survived through the more favourable conditions while it became extinct on the continent; or, a few individuals of a dominant species reached the island, where, owing to the absence of competition, they rapidly increased till the island became fully stocked with the unchanged species. Then (and then only) survival of the fittest would begin to act, and the differences of food and climate, with the different kinds of enemies, would render some modifications of structure, form, or colour advantageous, and thus a new species would be formed by adaptation from the old one in almost exactly the same way as on the continent. In both these cases recognition-characters, to aid in the prevention of intercrossing, would be developed by natural selection. But if insular species have usually been formed by a few individuals somewhat different from the type having first reached the island and thereafter preserved their peculiarities—as alleged by Romanes and others—there is no reason why any distinctive and stable form of coloration or marking should have been developed, since there would be no similar species from which it
1 Darwinism, pp. 174–180.
would need to be differentiated. Neither is the small amount of divergence that usually prevails between the mean of a few individuals taken at random, such as might have accidentally reached an island, and the average type of the species, at all comparable with the well-marked characters that usually distinguish insular forms, and there is nothing in mere isolation without selection which can increase the difference. As examples we may refer to the many peculiar species of butterflies and birds found in the various islands of the West Indian and Malayan Archipelagoes, which are quite as distinct from each other as are allied continental species, and which exhibit all the characteristics of species which have been fully differentiated by natural selection.1
1 As this point is still continually misunderstood by writers of considerable ability, it will be well to give here the substance of a reply I have made to their objections in Nature of January 12th, 1899.
The late Dr. St. George Mivart (in the Journ. Linn. Soc. Zoology, No. 172,) gives numerous cases of species of Lories peculiar to various Papuan or Pacific Islands, which differ in some details of coloration from the nearest allied species in other islands. He argues, as Captain Hutton had previously argued in the exactly parallel case of the fruit-pigeons of the genus Ptilopus, that these various specific markings cannot be useful as "recognition-marks" because the colour and marking of each of these genera is so very distinct from those of all other birds inhabiting the same island, there being usually only one species of a genus in each island. This conclusion is, however, not justified by the whole circumstances of the case, nor by the fundamental principle of organic evolution (which both these writers admit)—that each species has actually been developed from some allied species, living or extinct. Let us consider therefore what are the actual conditions of the problem. It is clear that each species of Lory or Fruit-pigeon now found isolated in any of the islands must (if evolution be admitted) have originated by modification from some parent species. The modification must have occurred in one or other of the modes suggested in the text, and the only mode that implies any special difficulty is when the species originated in the island where it is now found by the modification of some other species which accidentally reached the island. Let us therefore follow the process of change step by step.
The first thing that happens on the introduction of a few individuals of a species new to an island which is well suited to it, and where there are no other enemies than those to which it is already adapted, is its rapid increase in numbers till the island is fully stocked. That this happens we know from the increase of the rabbit in Australia, New Zealand, and Porto Santo, the sparrow in the United States, and numerous other cases. But as soon as the island is fully stocked a number equal to the annual increase must die off every year, and these will inevitably be, on the whole, those which are least fitted to the new conditions, whether as young or adult, and thus natural selection begins to act in the production of a new form suited to the new conditions. If the conditions are identical with those of the former home of the species there will be no change; but if there are differences of conditions, either climatal, or in food supply, or organic, as will in most cases be sure to exist, then there will set in a gradual adaptation to these new conditions. Here arises the circumstance that my critics have overlooked, that a modification of one species into another must have occurred in every case, and during that modification the need for some "recognition-marks" to aid in checking the intercrossing of the incipient new species with the parent form will be of the first importance. All variations in colour or marking will be selected for this purpose, and thus will be produced those external specific characters which are so marked a characteristic of these insular forms. When the change has been completed and the parent form has died out, these special and characteristic markings or coloration will appear to us to be useless. But they were essential to the development of the new species, and having become fixed by long continued selection they now remain as a constant specific character.
Characteristics of Species due to Natural Selection.
The sketch now given of the usual mode of formation of new species under natural selection, leads to the conclusion that every species (of the higher animals at all events) will usually possess at least three peculiarities: in the first place, it must exhibit some difference of structure or function adapting it to new conditions; secondly, some distinction of colour, form, or peculiar ornament serving as distinctive recognition marks; and, thirdly, the physiological peculiarity of some amount of infertility when crossed with allied species. The first two constitute its "specific characters." But if we consider that every species in the long line of its ancestry must have had similar, but somewhat different specific characters, adapting it to the peculiar conditions of its environment and distinguishing it from its nearest allies; that some of these characters, when generally useful, have persisted, and now constitute generic or family characters; that others have been again and again modified so as to adapt them to new and sometimes quite different conditions; and that others again, becoming useless, persist when quite harmless or
VOL. I. C C
remain in a more or less unstable or rudimentary condition; and when we further consider that many genera and families extend far back into geological time and must have originated in the midst of a physical and biological environment very different from that which now prevails, we shall dimly understand how complex are the forces and processes which have led to the assemblage of characters now presented by each organism, and how difficult it must be to determine positively that any one of these characters is not, nor ever has been, useful to its possessor. Yet this is what is done by those writers who maintain, as did the late Mr. Romanes, that the majority of specific characters are not and never have been useful, but have arisen through definite variation under the influence of definite causes, and, when neither useful nor hurtful, persist and constitute the main external differences which we observe between species and species. This theory, which, although to some extent held by Darwin himself, I consider to be wholly erroneous, we will now proceed to discuss.
It may be well first to dispose of a point, made much of by Mr. Romanes, that I do not urge utility as a characteristic either of varieties or of genera and higher groups, and that it is therefore illogical to claim it for species. But this is a misapprehension, since I do claim that when varieties are constant, are hereditary, and occupy a definite area, and are therefore what Darwin termed "incipient species," the characteristics which distinguish them from the parent species are, to some extent, adaptive and useful, and will become fully so when the variety becomes a fully differentiated species. And as to genera and families, it is obvious that every one of their distinguishing characters was once a specific character, since genera are merely groups of species, all of which were derived from one parent species, and which have become more or less isolated by the extinction of intermediate forms. Families are, in the same way, derived from a single genus and ultimately from a single species, and the same reasoning applies to them. The reason why my argument on this question has been limited to species is,
because the whole problem is included in that of species; it is in them that the process and laws of development can be best studied free from many of those complexities of modification and survival of disused and partially aborted parts and organs which often constitute generic or family characters. If every one of the new characters or new combinations of characters which arise when a new species becomes differentiated from its parent-form,—if every one of these is adaptive and utilitarian, then no higher groups can possess characters other than those which were once adaptive, since genera and families can never acquire new characters except through every one of their component species acquiring those characters. The problem as exhibited in species includes therefore the problem in all higher groups.
Characters fixed by Natural Selection must be Useful.
I have already set forth in some detail the argument for utility founded on the fact of the continuous progress of the discovery of utilities with the continuous growth of our knowledge of the life-histories and inter-relations of plants and animals.1 I will therefore now devote more special attention to the fundamental argument, that whereas every modification of a species which arises under the influence of natural selection must, from the very nature of its origin, be useful to the new form, no other agency has been shown to exist capable of producing non-utilitarian characters in every individual constituting a species, neither more nor less. Now the general cause which is adduced as being able to do this is stated by Darwin in the following passages, which are quoted by Mr. Romanes as expressing his own views:—
"There must be some efficient cause for each slight individual difference, as well as for more strongly marked variations which occasionally arise; and if the unknown cause were to act persistently, it is almost certain that all the individuals of the species would be similarly modified" (Origin of Species, p. 171).
1 Darwinism, pp. 131–142.
C C 2
Again, after referring to cleistogamic flowers and degraded parasitic animals, he says:—
"We are ignorant of the exciting cause of the above specified modifications; but if the unknown cause were to act almost uniformly for a length of time, we may infer that the result would be almost uniform; and in this case all the individuals of the species would be modified in the same manner" (Origin, p. 175).1
Now these passages, merely as stating a possibility or a probability, appear to me to be wanting both as regards logic and in the absence of any appeal to the actual facts of variation. For the argument is, briefly, that the same causes will always produce the same or closely similar results. But this is only true when the same causes act upon identical materials and under identical conditions. Now the very foundation of the Darwinian theory is, that the materials—the individuals of a species—are not identical, but that they vary indefinitely and in many directions even under closely similar conditions. How then can any external or internal causes produce an identical result—a definite new variation—in all the individuals of a species, born as they are of varying parents, of different ages, and subject to ever fluctuating conditions? It seems to me, therefore, that the a priori probabilities are all against Darwin's supposition.
Now let us see how far the facts of variation give any support to the theory of useless specific characters. If there is one thing better established than another it is that the individual variations which are constantly occurring in all common species are indefinite in their character and very unequal in their amount. Some species are much more variable than others, and Darwin has shown reasons for believing that any change of conditions induces variability, but not that it causes definite variations. The two things are radically distinct. So far as I am aware, no evidence has been adduced of any special con-
1 In my Darwinism, p. 141, I have stated my opinion that Darwin did not believe in the production of useless characters in all the individuals of a species. I had overlooked the passages quoted by Mr. Romanes and given above, which certainly show that he did believe it.
ditions which have produced a definite variation in the whole offspring of all the individuals subjected to it. But it must do more than this. For it must produce a variation so exceptionally stable that it constantly recurs in all the offspring of successive generations, even though those offspring are subjected to considerable change of conditions, as are the individuals of all species except the rarest or the most local. Only with such constancy and stability of inheritance could a useless character become fixed in every individual of a species, which it must be to be a "specific" character. It must, therefore, from the very first have been invariable. But this feature of invariability without selection has not been found to characterize any variation, whether occurring among wild or domesticated organisms. Such an occurrence would necessarily have forced itself upon the attention of breeders and horticulturists. For if the theory is true that the majority of specific characters are of this useless kind, their occurrence as permanent and unchangeable variations must be a common phenomenon, and we ought to find that foreign plants when first cultivated very often present new characters, not sporadically but appearing in every individual, and which cannot be got rid of, since they do not vary, and selection would therefore be powerless to eliminate them. Has any indication of a phenomenon of this kind ever been noted?
Causes Alleged to Produce Useless Specific Characters.
Let us come now to the actual causes said to produce useless specific characters. According to Mr. Romanes they are five in number: Climate, Food, Sexual Selection, Isolation, and Laws of Growth. Let us consider how these are known to act or are alleged to act. Climate and Food undoubtedly produce modification in the individual, but it has not yet been proved that these modifications are hereditary. If this could be proved the whole discussion on the heredity of acquired characters would be settled in the affirmative. The supposed proof that these causes produce definite changes which are
hereditary is derived from the fact that there is often a simultaneous change in the colours of many animals, or in the form or texture of the foliage of many plants, in different parts of the area they occupy which are characterized by differences of climate. But in every case these changes can be interpreted as adaptations for protection in the case of the animals, and as either adaptations or individual non-hereditary modifications in the case of the plants. The firm belief that such individual characters were usually, if not always, inherited, led to some looseness in Darwin's reasoning on this point, and still more so in that of most modern upholders of the theory.
The next alleged cause, Sexual Selection, whether we limit it, as I do, to the struggles of the males, leading to the development of weapons and defensive armour, or with Darwin extend it to the choice by the females of the more ornamental males, thus leading to the development of decorative plumes, &c., is really a form of natural selection, and sexual characters are therefore useful characters. It is true that, from my point of view, male distinctive colour and ornament have not this particular use; and Mr. Romanes makes a good point against me when he says that in imputing their origin and development to the surlpus vitality and energy of the male I give away my case, since I admit that useless specific characters may be developed independently of natural selection. This is owing to my having omitted to lay special stress on the specific part of each ornament being really a "recognition mark," and therefore essential both to the first production and subsequent well-being of every species. In the summary of my argument (Darwinism, p. 298) I have adduced the need of recognition as the cause of specific specialisation of colour, but in the body of my discussion as to sexual ornaments I have not referred to it, and this omission greatly weakens my argument. I should have said that the accessory plumes and other ornaments originate at points of great nervous and muscular excitation, and are developed through surplus energy; and that from their first appearance, they were utilized for purposes
of recognition, which explains both their comparative stability in each species and their distinctness in allied forms.1
The next alleged cause, Isolation, I do not admit to be a vera causa at all, for reasons already given. It is, at most, an aid to the differentiation of new species by natural selection.
The last alleged cause, the Laws of Growth, can never, of itself, account for specific characters, but only for those structural and histological peculiarities of organisms which characterise the higher groups such as classes and sometimes perhaps orders and families; and even these must always, when they first originated, have had a utilitarian character, since it is almost impossible to conceive that the details of structure of the various tissues or organs produced under the action of these laws were absolutely indifferent to the well-being of the organism.
If, then, we admit, as I do admit, that certain growths appendages, or markings, which are of no use to the organism, do occasionally appear, no agency has been adduced which could, first, cause these useless characters to appear in every individual of a species, and then totally cease to appear whenever any portion of this species is selected and slightly modified so as to occupy a new place in nature or to save itself from extinction by some new enemy. Whenever useless characters are said to be "specific," it seems to be forgotten that one species has always passed continuously into another by a process of normal individual variation and survival of the fittest. There is no chasm in such a process, no sudden transition
1 Since this chapter was written I have carefully studied Professor Weismann's new theory of "Germinal Selection," which seems to me to have a high degree of probability, and which, if true, enables us to explain two phenomena which have not hitherto been fully explicable. These are (1) the complete or almost complete disappearance of many characters which have become useless; and (2) the development of secondary sexual characters far beyond the point of utility as recognition-marks, and, apparently, up to the extreme point of incipient hurtfulness. It thus furnishes the one link necessary in the chain of argument proving that these secondary sexual characters are explicable without calling in the very problematical agency of female choice.
from one creature to another of a different nature. The transition is by a purely normal and almost imperceptible process of adaptation to new conditions, and in itself furnishes no reason whatever why any useless character, if it had constantly reappeared in the countless millions of individuals during all the millions of generations of the duration of the species, should at once disappear, or be replaced by some new character equally universal, equally invariable, and equally useless.
I strongly urge, therefore, that the general causes suggested by Darwin as possibly leading to the production of useless specific characters, as well as the more special causes enumerated by Mr. Romanes, do not apply to the actual facts of variation and heredity so far as they are yet known to us; and further, that no attempt has been made to show, even hypothetically, how, through the action of known causes, such characters, when they do arise, can become, first, extended to every individual of a species, and then be totally obliterated as regards any portion of the species which may become modified so as to constitute a new species. Useful characters thus strictly limited are the necessary and logical results of modification through survival of the fittest. No agency has been shown to exist capable of producing useless characters similarly limited. And as it is beyond the powers of human reason to know absolutely that any characters so limited as to be really specific are and always have been useless, it is both unscientific and illogical to postulate such characters as being present in all or many species, and therefore as constituting an essential characteristic feature of specific forms.
The preceding discussion may, I hope, be considered sufficient to show that useless specific characters, if they exist, can only be the result of some comparatively rare and exceptional conditions, and that they certainly are not, as has been alleged, a general characteristic of species; but it may be as well to notice a few of the special cases which have been adduced by Mr. Romanes and others as examples of their existence or as illustrating their formation.
Special Cases Discussed.
The Niata cattle of South America, which have strangely upturned jaws, are said to breed very true and to form a definite well-marked race which, if the character were not injurious but simply indifferent, might lead to the formation of a species defined by this useless specific character. The short-legged Ancon sheep, and the six-toed cats, are other examples of such remarkable abnormalities or sports which have the curious property of being strongly hereditary, and yet, apparently, of never leading to the formation of new species. Almost all students of evolution now admit that "sports" or large and sudden divergencies from the specific type are not the materials from which new species have been formed, the reason being that they are extremely rare occurrences; and when any such "sport" appeared in a species, the individual presenting it would either be avoided by its fellows and leave no offspring, or by repeated crossings with the normal type the sport would disappear. We may, no doubt, imagine conditions under which a sport of this kind, once appearing in both sexes, might lead to the formation of a breed and ultimately of a species; but the combination of conditions requisite to bring this about is so improbable that we can only look upon it as a bare possibility. But the question we are discussing is not whether, under certain very rare and exceptional conditions, a few species may possibly be formed which are distinguished only by altogether useless characters, but whether such characters are common in the majority of species and, to use Mr. Romanes' words, exist in "enormous numbers." The case of abnormal sports or monstrosities such as those here referred to can certainly not be adduced as giving any support to this view.
The next case, that of the Porto Santo rabbits, is held by Mr. Romanes to prove that the constant characters which distinguished them from common rabbits were only results of the action of peculiar conditions on individuals, and were not produced by natural selection. He arrives
at this conclusion from the fact that one of the two which died at the Zoological Gardens after four years' captivity was sent to Darwin, who found that the special colouring that distinguished the breed—the absence of black on the tail and ear-tips and the reddish colour on the back—had almost disappeared, and that the whole colouring was very little different from that of the common wild rabbit. Hence Mr. Romanes concludes that other wild species may be really only climatal forms, and their peculiar characters be non-adaptive. But no mention is made of the remarkably small size of these rabbits, which were only about half the weight of the common wild species and which looked no larger than average rats. If this also were a result of the action on the individuals of scanty food or a peculiar climate, it would have rapidly disappeared with ample food at the Zoological Gardens; and neither in this point nor in the peculiar form of the posterior end of the skull and interparietal bone, which was so distinct that Darwin figured it (see Animals and Plants under Domestication, i. p. 118), did he note any difference in the dead animal. It seems probable, therefore, that the colour-peculiarities of the Porto Santo rabbits were due to a change of tint of the longer hairs which may have been lost during the illness which led to the animal's death. And as we have no information as to the supposed change having been progressive during the four years of confinement, or that it affected the second specimen, no such conclusion as that drawn by Mr. Romanes can be held to be established. But, strange to say, Mr. Romanes did not notice that his own interpretation of this case is directly opposed to the theory of the heredity of acquired characters which he was disposed to favour. For these rabbits had lived in Porto Santo under identical conditions for at least 470 years, that is, for at least 470 generations. And yet the acquired characters had not become fixed in all these years; but, when subjected to new conditions, the very individuals brought from Porto Santo changed back, according to Mr. Romanes, to the original type of the common wild rabbit. If this were so, surely there never was, or hardly can ever be, a more
conclusive proof of the non-heredity of acquired characters. He declared that this case "definitely and conclusively" overturns the assumption of natural selection having acted in this case, but he omitted to state that it also (if his interpretation is correct) definitely and conclusively overturns the assumption of the heredity of acquired characters, which in the same volume he has elaborately discussed.
The only other case of much importance is that of changes of colour said to be directly caused by changes of climate, and especially by darkness in cave-animals. In this latter case it is declared by Mr. Romanes that the loss of colour cannot be of any use and cannot have been caused by natural selection. It is, therefore, an example of a useless character occurring in all the individuals of many unconnected species. In the case of the Proteus however, it is stated that when subjected to the action of light in confinement, the skin becomes dark, showing that the character is in some degree an individual one, due probably to deficiency of nutrition or, partially, to the need of light for the secretion of the pigment. The whiteness is here not a specific character. And if, in other cases, it is permanent and specific, it may have had a very obvious use in the early stages of the modification of a cave-fauna. For if any animals were isolated in caverns which were not totally dark, the light tints would be important as recognition marks, enabling the sexes to find each other; and when, at a later period, the species spread into the parts which were totally dark, there would be no cause leading to a return of the positive colour, especially as all cave-animals subjected to total darkness must at first have been in great danger of extinction from deficiency of food, and there would thus be no surplus nourishment available for the production of pigments.
Several biological friends with whom I have discussed this question, while agreeing that the majority of specific characters are useful, have suggested that useless characters may have been produced in some such manner as the following. If some useless character appears as a
variation in some individuals of exceptional vigour, it may increase by interbreeding, and its repeated production being perhaps favoured by some local conditions, it may come to form a marked local variety. Now, if the conditions become unfavourable to the species in the area occupied by the type, this may in course of time become extinct, and the variety distinguished by the altogether useless character will remain as the only representative of the species. It may be admitted that such a mode of origin of a non-utilitarian specific character is conceivable, but whether it ever actually occurs in nature may be doubted; while if it does occur, it must be owing to so rare a combination of circumstances that it can produce no such general prevalence of useless specific characters as is claimed by the advocates of that theory.1
In order to ascertain whether the immediate antecedent to such a mode of species-formation as is suggested is at all common, and thinking that British flowering plants offer the best materials for its detection, I put the case to two experienced British botanists as follows:—Are there any examples within your knowledge of well-marked varieties (not mere individual states due to local conditions) which occupy a considerable area to the exclusion of the parent species, and which do not occupy any area, or only a very small one, with the type? Each of them suggested several species which seemed to answer to the conditions, but on further consideration it appeared that they did not do so, and we were finally reduced to a single case, that of one of the species of Rubus, a genus which most botanists will regard as a very unsafe one to draw any conclusions from. Rubus radula, Weihe, is said to be abundant in the Midland parts of England, but in the Southern and South-western counties to be replaced by the variety anglicanus of W. M. Rogers, the type never having been found in the area occupied by this variety. If this is the case, and the two forms, said to be easily recognizable, really occupy distinct areas and nowhere
1 If, however, the variation is preserved because it occurs in exceptionally vigorous individuals, it is correlated with a character which is useful.
overlap, or very slightly so, then we have the condition precedent to the formation of a species by the extinction of the type, thus leaving the variety to represent the species. Of course in this case we do not know that the characters which distinguish the variety are useless; but if they are so, and if the variety should possess some superior vigour of constitution or other useful peculiarity which enables it to survive when the type dies out, we should have an illustration of one mode in which useless specific characters may possibly have arisen.
The inquiry is interesting, however, because it brings to light the rather unexpected fact, that fixed varieties of plants occupying considerable areas to the exclusion of the type are not common, and, perhaps, in our island do not exist. And should they be found to occur more frequently in other countries—as varieties of birds, mammals, and reptiles do occur in separate areas in North America they may be usually explained as adaptations to very different climatic conditions, in which case the distinguishing characters will be utilitarian, and the local varieties will be really incipient species.
Conclusions from the Inquiry.
The preceding inquiry leads us to certain very definite conclusions. In the first place, we see that species, which have been differentiated as such by the laws of variation and survival of the fittest, must be characterized by certain peculiarities whereby they have obtained an advantage in the struggle with their fellows. These peculiarities constitute their "specific characters," and these must be useful. As this applies also to every species in the direct line of descent, the characters which are sectional or generic must also, at the time of their origin, have been useful.
In the second place, although non-utilitarian characters do undoubtedly appear in the normal course of variation, no agency has yet been detected adequate to the extension of these useless peculiarities to all the individuals which constitute a species, and, further, to prevent their
extension to any of the varieties which are destined to become new species. Unless the power in question can have this twofold effect it cannot lead, except by accident, to the production of useless specific characters.
Under conceivable conditions, however, it is possible that certain useless characteristics may become limited to the individuals of a single species. But what we know of the modes of variation and the distribution of varieties indicates that, if at any time so produced, they must be altogether exceptional and of the nature of chance products; and that they cannot possibly constitute such a general characteristic of species as has been suggested.
Our final conclusion is that, whether we can discover their use or no, there is an overwhelming probability in favour of the statement that every truly specific character is or has been useful, or, if not itself useful, is or has been strictly correlated with some useful character.
CHAPTER XIX
THE POLYNESIANS AND THEIR MIGRATIONS
THE origin of the various races of the islands of the Pacific has always been one of the most difficult problems for the believers in the unity and the recent origin of man. Their diversity of physical features, of civilization, and of language, the absence of any continental races to which they could be affiliated, and the wide spaces of ocean over which they are distributed, have hitherto seemed to indicate that their origin dates from a period so remote that we cannot hope to determine it with any approach to certainty.
M. Quatrefages,1 however, evidently believes he has completely solved this difficult problem, as far as regards the most important of the Pacific races, the Polynesians. He very properly limits this term to the brown races spread over a wide area from the Sandwich Islands in the north to New Zealand on the south, and from Easter Island on the east to the Tonga and Samoan groups on the west; but all speaking dialects of one well-marked language. Now what M. Quatrefages attempts to prove is, that these people are simply Malays, who migrated from some islands of the Malayan Archipelago (probably Bouru in the Moluccas), and have more or less intermingled with the races of Melanesia and Micronesia. His evidence to prove this is of two kinds;—first, he en-
1 Les Polynesiens et leurs Migrations, par M. de Quatrefages, Membre de l'Institut, Professeur au Museum. Paris, pp. 199, Arthur Bertrand, Editeur, 21, Rue Hautefeuille.
deavours to show that a migration did take place; secondly, that the Polynesians are in their physical, mental, and moral characteristics, a true Malayan race.
1. Migrations.—We find in M. Quatrefages' volume a very careful summary of all the native accounts of their migrations, and also of the involuntary migrations that have recently occurred. These, no doubt, prove that the Sandwich Islands and New Zealand have been peopled by emigrants from the Marquesas and Tahiti, and the fact of this emigration is confirmed by the independent evidence of language. It is proved, therefore, that the Polynesians have passed over immense spaces of ocean, in directions not especially favoured by winds or currents, and thus the difficulty of any migration from its mere distance is quite overcome. It is further shown that all the traditions point to the Samoan group and the Fiji Islands as the central points to which almost all Polynesians trace their origin. It is to be observed here that these are the largest of all the islands in the central Pacific inhabited by the Polynesian race, and it is these, therefore, that we should naturally expect to have sent out colonies to the smaller islands. So far we have the strongest corroboration of there having actually been a migration, in the fact of the community of language, and all the legends of these migrations speak of them as having been made by simple men, the natural ancestors of the existing Polynesians. But in the legend which refers the origin of the Samoans themselves to a migration from a large country "further west," we get into pure legend,—for the mythic Bouloton, whence the first inhabitants are said to have come from, is a spiritual and not a real country, and these inhabitants are believed to have been not men, but inferior gods. But the direct evidence, so far as it goes, of the earliest migration having been from the west, is by no means so clear as M. Quatrefages would have us believe; for one of the best authorities on the subject, Mr. W. P. Pritchard, who has spent his whole life in the Pacific, and from his long residence in the Feejee and Samoan Islands as British Consul, and his intimate
knowledge of the Polynesian languages, is well qualified to give an opinion on this matter, says it is quite the contrary. In his Polynesian Reminiscences, p. 402, he observes: "It is, however, remarkable that in all these many instances of authenticated driftings, the course of the drifted canoes has been from east to west before the prevailing trade winds, and not from west to east before the westerly winds;" during the prevalence of which he tells us the natives do not usually venture out on fishing or travelling expeditions.1
In this case, too, the corroborative proof by language completely fails, for though there is an undoubted Malay element in the Polynesian language, it is an element derived from the civilized Malay and Javanese tongues, not from those of the Moluccas, which are totally distinct. It is to be noted also that this Malay element in the language has all the character of a recent introduction, since the Malay words are hardly changed, except by the phonetic character of the language which has received them.
But even if the Malay formed a much larger portion of the Polynesian language than it does, this would not prove a community of race, unless the physical characters also in some degree corresponded. It is here that we find an absolute defect of all evidence bearing upon the point in question—the similarity of the Polynesians to any race speaking the Malay language. Almost the only evidence adduced by M. Quatrefages is as to the similarity of the brown race of Timor to those of Polynesia. But the Timorese are not Malays at all; they belong to that curious race which has close affinities to the Papuan in all
1 Later and more extensive inquiries have shown that accidental drifting of canoes from north or north-west to south-east does often occur, especially in the region of the northern tropic, where violent storms and hurricanes are not unfrequent, and it is when unexpectedly caught by such storms that canoes get carried far away from their course, and occasionally drifted to distant and unknown islands. Such was the immense double canoe with men, women, and children, which over a hundred and fifty years ago reached Rotuma from Tarawah in the Gilbert Islands. This was a journey of over a thousand miles in a direction south by east. Numerous other cases have been recorded in various Pacific Islands of canoes carried by storms from the north-west; and most anthropologists now believe that the Mahori race originally came from some part of the Asiatic continent or its adjacent islands.
VOL. I. D D
moral and physical characteristics except colour, and their languages are much further removed from the Malay than even the Polynesian itself. The resemblance physically of this race with the Polynesian proves absolutely nothing with regard to the Malay question.
Now let us compare the most important and thoroughly well-established physical and mental characteristics of the two races:—
| Polynesians. | Malays. |
| Tall, averaging— | Short, averaging— |
| 5 ft. 10 in … Wilkes (Samoans). | 5 ft. 4 in.or 5 ft. 6 in. |
| 6 ft.. Dupurey (Tahiti). | |
| Hair wavy, curly or frizzly. | Hair always straight. |
| Beard often full. | Beard almost wanting. |
| Face handsome, European type. | Face of Mongolian type. |
| Nose often aquiline. | Nose never aquiline. |
| Disposition active and joyous. | Disposition slow and morose. |
| Character open and frank. | Character eminently secretive. |
| Often erected stone edifices. | Never used stone for building. |
| Use double canoes. | Use single canoes. |
Taking the whole of these differences, they appear to indicate a radical diversity not to be overcome by any mere similarity of colour and some common words in language, which is all that really exists to prove identity of race. The one single fact of stature is conclusive against any such comparatively recent common origin as M. Quatrefages argues for. A race which averages 5 ft. 10 in., and has many men 6ft. 2 in. or more, can hardly have been derived at such a recent period as to have retained community of language from a race averaging 5 ft. 5 in. or 5 ft. 6 in., and among whom a single individual of 5 ft. 10 in. is rarely, perhaps never, found. Again, the hair of the Malay is of the true mongol type—black, coarse, and perfectly straight.
The least approach to wavy or curly hair is never found among the unmixed Malays. I cannot find evidence that the Polynesians ever have this character of hair, while it is not unfrequently as frizzly as the most decided Papuans, but that is no doubt due to intermixture. Again, the mental character of two races in a parallel state of civilization and inhabiting very similar countries, is surely of great importance; yet, what contrast can be greater than between the phlegmatic, suspicious, undemonstrative Malay, and the active, frank, and joyous Tahitian? Are we to throw down all these barriers of diversity for the sake of solving by main force a problem that is probably insoluble?
We ought not to expect that the beginnings of every race can be discovered within the short epoch of human history or tradition, and we have every reason to be suspicious of the theory that professes such a discovery. In the present case, the very erroneous views prevalent on the subject arise from two causes. One is the occurrence of a number of Malay words in the Polynesian language; the other, the similarity of the brown tint of the Malays and Polynesians, while they are separated by a group of people of a much darker colour. The similarity of tint has led many travellers in the one area to jump to the conclusion that the people of the other area, of which they have little knowledge, are the same race. It unfortunately happens that not a single traveller appears to be well acquainted with both races, and for that reason their opinions as to the similarity of the two should be received with great doubt. If, on the contrary, my account of the physical and mental characteristics of the Malays be taken as correct (and I resided among them for eight years), and if it be compared with that of the Polynesians given by Cook, and by recent travellers and missionaries, the differences will be seen to be so striking and radical, that all idea of their being the same race must be given up. In the case of the Malays in particular, much confusion has arisen from travellers having confounded with them the many peoples of distinct race which inhabit the eastern parts of the
D D 2
Malayan Archipelago, such as the Timorese, the mountaineers of Ceram and Gilolo, and of the small islands near New Guinea; and this mistake has been rendered excusable by the number of half-breeds between the two races to be found everywhere. Many of these people are, perhaps, allied to the Polynesians, but they are certainly not Malays, who are essentially a Mongol race, with many of the Mongol characteristics strongly marked. The Papuans of New Guinea form the extreme type of another and a widely different race, and all the evidence goes to show that in every characteristic except colour, the Polynesians are somewhat nearer to the Papuans than they are to the Malays, but that they are fundamentally distinct from both of them.
Modern Views as to the Affinities of the Polynesians.
Since the preceding pages were written, more than thirty years ago, a great advance has been made in our knowledge of the true affinities of the Mahoris or brown Polynesians. The supposed close relationship of their languages to the Malay has been shown to be an error, since they differ fundamentally both in phonetics and in grammatical structure. The Malayan words in the Mahori languages, once thought to be so important, have been shown to be not more than about four per cent. of the whole, and they belong mostly to the actual spoken languages of the western Malays and Javanese, not to those of the more eastern tribes in the Moluccas or the Philippines. This is easily explained by the fact of the two former languages being those of the wandering traders of the Archipelago, who would be most likely to come in contact with the Mahoris and thus introduce some words of their language.
Another factor of great importance which has led to a more accurate perception of the resemblances or differences of the races of distant lands, has been the great spread of photography during the last twenty or thirty years. So long as we were dependent on the hand-work of artists of every degree of talent, and often with such an amount of
personal equation or mannerism as to disguise some of the most important peculiarities of a race, and when their sketches, often hasty or taken from individuals who were not typical of their race, were still further modified by the copyist or engraver, no accurate comparison of types was possible. But with the advent of the photographer and of the process-plate this difficulty has disappeared, and we are able for the first time to make accurate comparisons. The chief difficulty that remains is due to the ignorance of the photographer, who is too often a passing traveller or tourist, and quite unable to distinguish between pure and mixed blood, or between individuals who are typical of the best qualities of their race and those who are altogether exceptional. This difficulty however tends to disappear with the multiplication of anthropological portraits, and especially when they are taken by intelligent residents who are well acquainted with the special characteristics of the people amongst whom they live. But even at the present time it is very difficult to get really good portraits of the best types of these interesting people.
Almost all travellers speak highly of the appearance and character of the brown Polynesians. Darwin, who only saw them at Tahiti, says:—"I was pleased with nothing so much as the inhabitants. There is a mildness in the expression of their countenances which at once banishes the idea of a savage; and an intelligence which shows that they are advancing in civilization." And after taking part in a meeting between the principal chiefs and Captain Fitzroy about compensation for injury done to an English ship in one of the most distant islands of the group, he says: "I cannot sufficiently express our surprise at the extreme good sense, the reasoning powers, moderation, candour, and prompt resolution, which were displayed on all sides." Hellwald, after referring to the deterioration since the days of Cook, due to the introduction of liquor and other vices of civilization, says:—" The natives of Tahiti are still a fine, well-proportioned people, tall and robust, with dark brown complexion, broad nose, slightly protruding lips, beautiful
teeth, black and mostly curling hair, but with slightly developed beard."
The two portraits here given of a young man and
FIG. 52.—MAN, TAHITI.
woman reproduced from Challenger photographs by permission of the Stationery Office, sufficiently show the general character of Tahitians at the present day.
In the Sandwich Islands, the Kanakas, as the natives
are called, seem to have suffered most from the injurious effects of a too rampant civilization, since from Cook's estimate of 300,000 for the population in his day they are
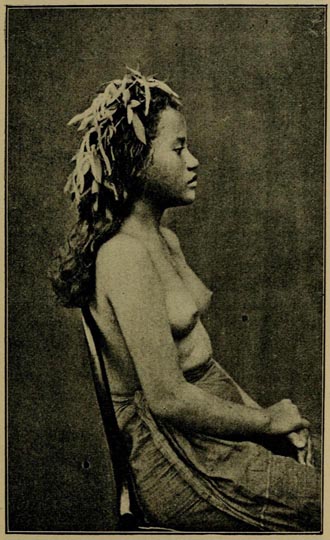
FIG. 53.—GIRL, TAHITI.
now reduced to a little over 30,000; while that most terrible of diseases, leprosy, is permeating all classes of the people, although quite unknown previous to 1840. It only became prevalent after 1860, about which time
vaccination was introduced by the missionaries, who in this matter were blindly followed by the people, and there is very strong evidence that in many countries it has been

FIG. 54.—MAN, HAWAII.
the means of spreading the disease.1 The Sandwich Islanders have been considered by most writers to be
1 For facts bearing on this question see The Recrudescence of Leprosy, by William Tebb (Swan Sonnenschein, 1893).
among the most intelligent and finest peoples of the Pacific, and it is a melancholy satire upon the supposed superiority of our civilization, that it should deteriorate

FIG. 55.—GIRL, HAWAII.
and destroy. The accompanying two photographs from the Challenger collection show us good types of Hawaiian men and women at a recent period.
It is, however, in the Friendly and Navigator's Island of the
western Pacific that the highest types of the Mahori race are still to be found, though these too have greatly diminished in numbers, while the intermixture with dark Fijians and low-class European traders have greatly deteriorated them. Admiral Erskine says:—"The men were a remarkably fine-looking set of people; among them were several above six feet high and of Herculean proportions. The manly beauty of the young men is very remarkable; one in particular, who had decked his hair with the flowers of the scarlet hibiscus, might have sat for an Antinous." The accompanying photographic print of a native of the Tonga Islands, kindly lent me by Mr. J. J. Lister of St. John's College, Cambridge, well illustrates this fine type. Mr. Lister informs me that the "sisi" or collar worn by this man was made of strips of hibiscus bark and ferns brought from Ena, one of the volcanic islands of the group not far from Tongatabou.
Lord George Campbell in his delightful book, Log-letters from the Challenger, speaks of these people in enthusiastic terms. "There are," he says, "no people in the world who strike one at first so much as these Friendly Islanders. Their clear light copper-brown coloured skins, their curly hair and good-humoured handsome faces, their tout-ensemble, formed a novel and splendid picture of the genus homo; and as far as physique and appearance go, they gave one certainly an impression of being a superior race to ours." And in character they are equally admirable. Cook declared them to be "liberal, brave, open, and candid, without either suspicion or treachery, cruelty or revenge." They are cheerful in disposition and fond of dancing and song; very cleanly in all their habits, so that Admiral Erskine remarks that "they carry their habits of cleanliness and decency to a higher point than the most fastidious of civilized nations." And again, that—"Their public meetings and discussions are carried on with a dignity and forbearance which Europeans never equal, while even in the heat of war thay have shown themselves amenable to the influences of reason and religion."
Their superiority is also shown in the status of women

FIG. 56.—NATIVE OF TONGA ISLANDS.
among them, who are treated as companions and equals, protected from all hard work and everything that might
impair their grace and beauty. The Mahori women employ themselves solely in household work, making mats and tapa cloth, plaiting ornamental baskets, and engaging in such outdoor employments as fruit-gathering and fishing, which in their delightful climate and in congenial companionship are relaxations or pastimes rather than labour. The Polynesians are kind and attentive to the sick and aged, and unlimited hospitality is everywhere practised; and, what is even more admirable and uncommon, the chiefs work as well as the common people, and think it a disgrace if they do not excel in all departments of labour.
The man and woman of Samoa here represented from photographs by Colonel Swinton are not of the highest type, but may be taken as average specimens. (Fig. 57 and 58)
How sad it is that all the more fertile islands inhabited by this most interesting race—a race that seems, under favourable conditions, capable of developing a true civilization higher than any the world can yet show, should be seized upon by the great nations which especially pride themselves upon their civilization, not for the purpose of helping on the development of this fine race which the world can ill afford to lose, but solely in their own interests, political, naval, or commercial. If we contrast this conduct and the national character it implies, with that of the Mahoris as depicted by unprejudiced persons from Captain Cook down to our own day, we shall be driven to the conclusion that in all the essentials of true civilization these uncultivated people are fully the equals—perhaps even the superiors of ourselves.
The Mahoris are Caucasians.
Keeping in view the mental and moral characteristics of the Mahori peoples as here briefly indicated, and considering carefully the physical characters shown by the photographs here reproduced, we shall not be surprised to find that the tendency of modern anthropologists is to class them as belonging to the same great primary division
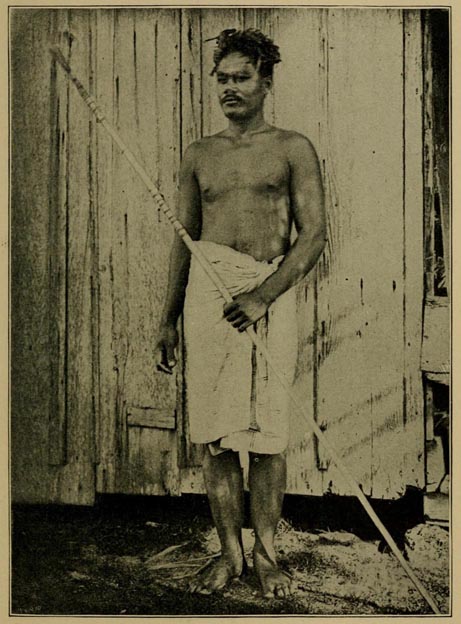
FIG. 57.—MAN OF SAMOA. (TAKEN IN 1893, COL. SWINTON).
of mankind as ourselves—the Caucasian. Professor A. H. Keane, speaking of the Polynesians, says:—"A line drawn athwart the Pacific from New Zealand through Fiji to

FIG. 58.—WOMAN OF SAMOA. (FEB. 1893. COL. SWINTON).
Hawaii will roughly cut off this domain from the rest of the Oceanic world, where all to the west is Melanesian, Papuan or mixed, while all to the east, the Maori of New
Zealand, some of the eastern Fijians, the Tongans, Samoans, Tahitians, Marquesans, Hawaiians, and Easter Islanders—constitute the purest and most interesting section of the Caucasic Indonesians. Their claim to belong to this connection can no longer be seriously questioned, since, as now firmly established, there have been from the remotest times both a dolicho-and a brachy-section of the Caucasic division. To the former section belong our Eastern Polynesians, who are mostly long-heads with remarkably regular features, often of a distinctly European stamp, and other characters of a pronouncedly Caucasic type."
Professor Keane goes on to discuss their language, which is not Aryan but Malayo-Polynesian, which had its roots on the Asiatic main-land, whence it spread over the oceanic world in pre-historic times. He thinks they reached their present home through Malaysia, a point which seems to me very doubtful, and which will be referred to again when we have considered the origin of the Australians, and the widely scattered fragments of low-type or barbarous Caucasic peoples in the remote East.
CHAPTER XX
NEW GUINEA AND ITS INHABITANTS
IMMEDIATELY north of Australia, and separated from it at Torres Straits by less than a hundred miles of sea, is the largest island on the globe—New Guinea, a country of surpassing interest, whether as regards its natural productions or its human inhabitants, but which remains to this day less known than any accessible portion of the earth's surface. Within the last thirty years considerable attention has been attracted towards it, by surveys which have completed our knowledge of its outline and dimensions, by the settlement of English missionaries on its southern coasts, by the explorations of several European naturalists, and by the visits of Australian miners attracted by the alleged discovery of gold in the sands of its rivers. From these various sources there has resulted a somewhat sudden increase in our still scanty knowledge of this hitherto unknown land; and we therefore propose to give a general sketch of the island and of the peculiar forms of life that inhabit it, and to discuss briefly some of the interesting problems connected with its indigenous races.
It has hitherto been the custom of geographers to give the palm to Borneo as the largest island in the world, but this is decidedly an error. A careful measurement, founded on the most recent maps, shows that New Guinea is considerably the larger, and must for the future be accorded the first place. In shape this island differs greatly from Borneo, being irregular and much extended in a N.N.W. and S.S.E. direction, so that its greatest length is little
short of 1,500 miles, a distance as great as the whole width of Australia from Adelaide to Port Darwin, or of Europe from London to Constantinople. Its greatest width is 410 miles; and, omitting the great peninsulas which form its two extremities, the central mass is about 700 miles long, with an average width of 320 miles, a country about the size of the Austrian Empire, and, with the exception of the course of one large river, an absolute blank upon our maps.
This almost total ignorance is the more remarkable, when we consider how long the country has been known, and how frequently its shores have been visited. It was discovered in 1511, even earlier than Australia; and from that time Spanish, Portuguese, Dutch, French, and English vessels have continually passed along its coasts. Most of our early navigators—Forrest, Dampier, and Cook—visited New Guinea, and have given us some account of its inhabitants; while, more recently, many exploring and surveying ships—the Coquille and Astrolabe, under French; the Rattlesnake, Fly, and Basilisk, under English; the Triton and Etna under Dutch commanders, have added to our store of information. Among private naturalists and explorers, the present writer was the first to reside some months in New Guinea in 1858; since which time Dr. Miklucho Maclay, a Russian; Dr. Beccari and Signor D'Albertis, Italians; Dr. A. B. Meyer, a German; Mr. Octavius C. Stone, and several English missionaries, have all made important explorations and added much to our knowledge of the natural productions of the island and of the tribes residing on or near its coasts.
From these various sources we have obtained a tolerable knowledge of the outside margin of the country, but never extending more than twenty miles inland, except in the case of the Fly river, which Signor D'Albertis ascended for nearly 500 miles, reaching a point somewhat beyond the centre of the island. The north-western and south-western peninsulas of New Guinea are the best known portions, and both seem to be mountainous throughout. In the north, Mount Arfak, a little beyond Dorey Harbour, is from 8,000 to 10,000 feet high, while in the south-east
VOL. I. E E
the Owen Stanley range has several peaks which reach elevations of from 10,000 to 13,000 feet. The Charles Louis mountains, commencing near the south coast, east of Triton Bay, appear to run far in a south-easterly direction, and their summits are believed to be snow-clad, and are probably at least 18,000 feet high. If they continue eastward in the same general direction, they would pass about 100 miles to the north of D'Albertis' furthest point on the Fly river, and perhaps form a great curve till they merge in the Owen Stanley range in the south-east. This, however, is mere conjecture, for throughout the whole course of the Fly river the land was low, and only on one occasion were high mountains seen to the north-west. Combining this with the fact that for a length of nearly 700 miles the south coast of New Guinea is low and swampy with no high land anywhere visible, we are led to conclude that there is probably a continuous range of lofty mountains towards the north, while the south consists of wide alluvial tracts and of slightly elevated inland plains. This part of the island would thus somewhat resemble Sumatra turned round, but with higher mountains, which are probably not volcanic, and with a considerably greater width of land.
Although the Fly river penetrates so far into the interior, its size and depth in its upper portion are by no means what we should expect in a stream fed by a lofty mountain range close to the equator. It is, therefore, almost certain that larger rivers exist further west; while another large river certainly flows northward, having its mouth in a delta at the eastern extremity of Geelvink Bay. Until these rivers are explored, and, at least, the lower slopes of the hills ascended, we cannot be said to have much real knowledge of the interior of New Guinea.
Situated close to the equator, and extending only eleven degrees south of it, the climate of New Guinea is hot and uniform, and the rains abundant; leading here, as elsewhere in similar situations, to the growth of a luxuriant forest vegetation, which clothes hill and valley with an ever-verdant mantle. Only on the coasts nearest to Australia, and probably influenced by the dry winds
from that continent, are there any open or thinly wooded spaces, and here alone do we find some approach to the Australian type of vegetation in the occurrence of numerous eucalypti and acacias. Everywhere else, however, even in the extreme south-east peninsula and adjacent islands, the vegetation is essentially Malayan; but Dr. Beccari, who collected plants extensively in the north-western peninsula and its islands, was disappointed, both as regards its variety and novelty. On the Arfak mountains, however, he found a very interesting subalpine or temperate flora, consisting of araucarias, rhododendrons, vacciniums, umbelliferæ, and the Antarctic genus Drimys. The forests of New Guinea are everywhere grand and luxuriant, rivalling those of Borneo, and of Brazil in the beauty of their forms of vegetable life; and we cannot consider the collections yet made as affording more than very imperfect samples of the treasures they contain.
Mammals of New Guinea.
The animal life of this great island is better known and is perhaps more interesting. Its terrestrial mammalia are, however, singularly few, and, with the exception of a peculiar kind of wild pig, all belong to the marsupial tribe or the still lower monotremes of Australia. The tigers, apes, and buffaloes described in the fictitious travels of Captain Lawson, are here as much out of their real place as they would be in the Highlands of Scotland; while the tracks of large animals, supposed to be rhinoceros or wild cattle, actually discovered by recent travellers, are now ascertained to be those of the cassowary, which, so far as we yet know, is the largest land-animal of New Guinea. Large birds were also seen and heard, whose spread of wing was estimated at sixteen or twenty feet, and which beat the air with a sound compared to the puff of a locomotive; but these are found to be only a well-known hornbill of very moderate dimensions. In place of these myths, however, we have some very interesting realities, the most remarkable, perhaps, being the tree-climbing kangaroos of rather large size, which, although but slightly
E E 2
different in external form from the jumping ground-kangaroos of Australia, hop about among the larger branches of trees, on the leaves of which they feed. They have a bushy tail, with somewhat shorter hind legs and more curved claws than their allies; and they afford a
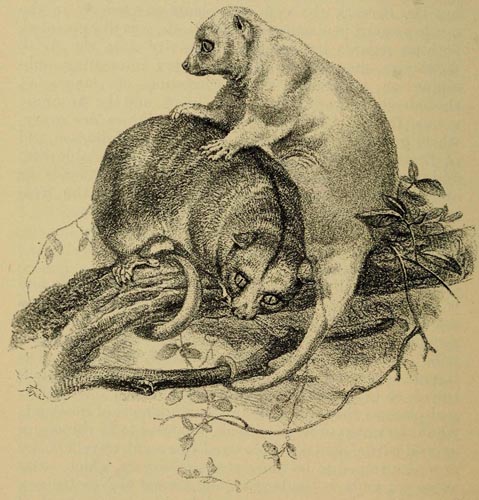
FIG. 59.—Cuscus orientalis (MALE AND FEMALE).
curious example of the adaptation of an animal to new conditions of life very different from those for which its general form and structure seem to fit it. Such a modification may, perhaps, be traced to a somewhat recent separation of Australia and New Guinea, when the kangaroos which remained in the latter country, not find-
ing a sufficiency of herbage for their support in the dense forests, began to feed upon leaves, and ultimately became adapted, with as little change as possible, to a truly arboreal life. The entire absence of beasts of prey would favour this adaptation, as the coincident acquisition of
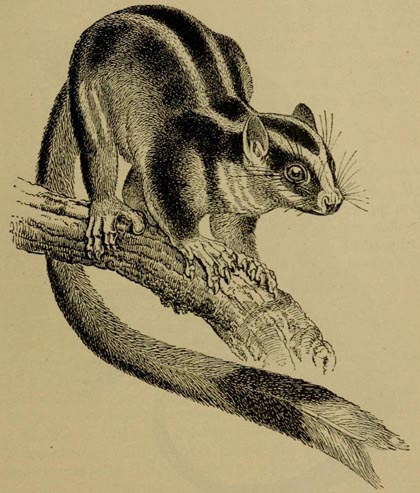
FIG. 60.—Dactylopsila trivirgata (ONE-THIRD NATURAL SIZE).
swiftness of motion or powers of concealment are thus rendered unnecessary; and the tree-kangaroo accordingly remains a slow-moving creature, just able to get its own living, but in all probability quite unable to cope either with enemies or competitors. (See figure at top of plate at p. 424).
Besides several species of Cuscus or Phalangers, one of the commoner species being that figured on p. 420, there are several allied genera of marsupials, among them the pretty little animal represented on p. 421, which has a rudimentary lateral membrane and thus forms a link

FIG. 61.—Distæchurus pinnatus (TWO-THIRDS NATURAL SIZE).
between the flying opossums of the genus Petaurus which are common in New Guinea, and Cuscus. It has also a very long tail, the hair spreading out bilaterally towards the tip, and this indicates that it probably does pass obliquely through the air from a high tree to a low one;
partially supported by the dilated skin between the outstretched limbs, and the broad tail. According to Mr. Semon it feeds on ants and other insects.
Another considerably smaller animal belonging to the same family is also figured (p. 422), and well shows the large grasping feet and hands characteristic of so many of the arboreal marsupials and seen also in the American opossums. Mr. Richard Semon, to whom I am indebted for the use of these two figures, had this creature alive for a few days, but found it to be very fierce and intractable. A curious form of the lowest known mammals, allied to the Echidna of Australia is also found in New Guinea.
Papuan Birds.
The birds, like the mammalia are mostly of Australian types, but nevertheless present many peculiarities. Most celebrated of all are the Birds of Paradise, forming a distinct family, containing more than twenty-five different species, all confined to this island and the immediately surrounding lands. These singular birds are really allied to our crows and magpies, but are remarkable for their special and varied developments of plumage. In most cases tufts of feathers spring from the sides of the body or breast, forming fans, or shields, or long airy plumes of extreme beauty. Others have glossy mantles or arched shields over the back, strange crests on the head, or long and wire-like tail feathers. These varied appendages exhibit corresponding varieties of colour. The long trains of waving plumes are golden yellow or rich crimson, the breast-shields, mantles, and crests, are often of the most intense metallic blue or green, while the general body plumage is either a rich chocolate brown or deep velvety black. The magnificent twelve-wired Bird of Paradise is figured in the upper part of the plate on p. 424. The breast and upper-surface are nearly black, but in certain lights rich tints of purple or green become visible. The top of the head is velvety and of a purplish bronze, the back and shoulders a bronzy green, while the closed wings and tail are of the most brilliant violet purple, and all of an exquisite silky gloss. The mass of broad plumes on

TREE KANGAROO. FIG. 62. Charmosyna papuensis.
Seleucides alba.
Tanysiptera galatea.
Goura coronata,
the breast are bordered with glittering bands of emerald green; while the large tufts of fluffy plumes on the lower part of the body are of a rich buff yellow. This fades into white after the dried skins have been exposed to the light, hence the misleading specific name of Seleucides alba. On each side six of these fluffy plumes have the shaft elongated into curled black wires, about ten inches long and bent outwards. We have here one of the finest contrasts of colour and strangest developments of plumage even in this wonderful family. This species is found in the north-western peninsula of the great island.
All the birds of paradise are exceedingly active and vivacious, the males meeting together in rivalry to display their gorgeous plumage, while the female birds are always unornamented and are usually plain or positively dingy in their colouring. From an unknown antiquity the natives of New Guinea have been accustomed to preserve the skins of these beautiful birds, and barter them with the Malay traders, by whom they were universally known as "burong mati," or dead birds, because they had never seen them alive. As the natives used always to cut off the feet in order to preserve them more easily, the Malay and Chinese traders concluded that they had none; and all sorts of stories were told about their living continually on the wing, and being in fact birds of heaven, whence originated the names of "Birds of Paradise" and "Birds of the Sun," given them by the early Portuguese and Dutch writers. Down to 1760 the skins of these birds never reached Europe with feet attached to them, and the great Linnæus recorded the fact by naming the largest kind Paradisea apoda, or footless Bird of Paradise, a name by which it is still known among men of science. The natives also generally cut off the wings, so as to give greater prominence to the ornamental feathers; and this gives the birds an altogether different appearance from what they really possess in a living state, or when properly preserved.
By far the greater number of these birds, and those of the richest colours and most remarkable plumage, live on the mainland of New Guinea, and they are especially
abundant in the mountains of the north-western peninsula, where the Italian and German naturalists already referred to obtained fine specimens of all the known kinds. In the south-east a new species has been discovered, but only two or three sorts are found there near the coast; and as they are also in little variety in the lowland districts of the north-west, it becomes pretty certain that they are more especially mountain birds. We may therefore confidently expect that, when the great ranges of the interior are visited and explored by naturalists, other and perhaps still more wonderful species will be discovered. It is interesting to note that, with the exception of one very peculiar species discovered by myself in the Moluccas, all the Birds of Paradise are found within the hundred fathom line around New Guinea, and therefore on lands which have probably been connected with it at a comparatively recent period.
Since the preceding account was first published, collectors in south-eastern New Guinea, both German and British, have discovered a considerable number of new species of these birds in the mountainous interior. The majority of these are more or less closely allied to the types already known, though sometimes more richly coloured or with a greater development of plumage. Such are several new species of the genus Paradisea or true Birds of Paradise, and the fine new six-shafted Bird of Paradise (Parotia Carolæ). This last is from the mountains of German New Guinea, and is distinguished by the brilliant bands on the back of the head, which are of iridescent violet, blue and green, while the crown is deep orange. Broad curved bands of silvery white over the eyes meet at the base of the beak. The scaly metallic feathers of the breast are rosy-violet and green, while the great tufts of side-plumes are pure white in the upper portion, bright chestnut in the middle, shading to nearly black in the lowest portion. The photograph (Fig. 63) is taken from a coloured plate which illustrates the description by Dr. A. B. Meyer, of the Dresden Royal Museum. The old species, of which this is a more brilliant form, was described by Buffon more than a century ago, and is
figured and described in my Malay Archipelago (p. 430).
Much more remarkable, however, is another species described and figured by Dr. Meyer from the same part of the island. The general plumage of this bird is by no means so brilliant as usual in the family. The upper

FIG. 63.—KING CHARLES' BIRD OF PARADISE (Parotia Carolœ).
parts are velvety purple-black with olive green glosses, the crown and lateral crests and the throat are purple-black; the breast is ochre-yellow; and the black wing-feathers are margined towards the base with rusty red. But its most striking and altogether unique feature consists of the two long curved appendages which spring from the outer corner of each eye close to the tear-gland,
These are fourteen inches long, the bird itself from beak to tail being about eight inches. They can hardly be called feathers, since they consist only of a slender shaft from the outer side of which grow a series of sub-quadrangular thin horny lamellæ resembling the fronds of some ferns. These lamellæ are of a beautiful blue colour (called by Dr. Meyer "forget-me-not blue"), thus strongly

FIG. 64.—THE FERN-BEARING BIRD OF PARADISE (Pteridophora Alberti).
contrasting with the whole of the body colours, and constituting one of the most striking and beautiful ornaments to be found not only in this wonderful family but in the whole class of birds. Even in this rich and still largely unknown island we can hardly expect to discover anything that will surpass this bird in eccentricity and beauty. It has been named Pteridophora Alberti, after King Albert of Saxony. The photograph from the coloured plate shows the general character of the species.
Two other species are sufficiently beautiful and distinct to be here referred to, and, by the kindness of Dr. R. B. Sharpe and Messrs. H. Sotheran and Co., I am allowed to give a photographic reduction of the plates which illustrate the former gentleman's beautiful monograph of the family. The first is Rothschild's Bird of Paradise.

FIG. 65.—ROTHSCHILD'S BIRD OF PARADISE (Astrapia splendidissima).
This magnificent bird somewhat resembles in general form the old Paradisé Pié (Astrapia nigra) briefly described in my Malay Archipelago (p. 435), and, like it, has none of the greatly lengthened feathery ornaments which usually characterise the species of this family, but it far surpasses that species in its varied and brilliant colouring, which are very inadequately indicated in the
photograph taken from an uncoloured plate. The general ground colour is velvety black with rich metallic purple and green reflections on the back; the head and neck above show various shades of metallic green and violet, with the hinder portion rich emerald green. The curved bands on the throat are metallic green and blue, with some fiery crimson beneath the eye; the feathers on the sides of the breast are edged with metallic steel green. The tail is glossed with purple, the two centre feathers being largely creamy white. The total length of this bird is fifteen inches, and in its extreme brilliancy it resembles some of the gorgeous humming birds on a gigantic scale. It was found near the base of the Charles Louis mountains in Dutch New Guinea, almost opposite the Aru Islands.
The other species referred to above is almost as peculiar and extraordinary as the wonderful fern-bearing Bird of Paradise, while its general plumage is far more beautiful. This is the Prince Rudolph's Bird of Paradise from the Owen Stanley mountains in south-east New Guinea. An examination of the figure here given (Fig. 66) shows that this bird possesses a very full development of the characteristic breast and body plumes of the family, and these differ from all other known species in being of various shades of blue, instead of yellow orange or red as in the corresponding plumes of other species. The head, neck, and body both above and beneath are velvety black with a yellowish green metallic sheen on the head, the eyelashes being long, silky, and white; the nape is a cherry brown, and the back greenish blue black. The wings above are blue, the feathers edged with brighter blue; the large breast-shield has blue and green glosses on the upper portion, with a broad blue band across the lower part of it. The ornamental lateral plumes are in three series, the longer being rather rigid, of a bright ultramarine blue passing into lilac at the ends, the next shorter series are bright cobalt blue passing into bright greenish blue at the tips, while the broad coverts are chestnut brown near the breast, black at the sides. The tail is blue above with brighter edgings, the two very long shafts being black, with a light blue spot
on the spoon-shaped tip. The bill is yellowish white. The female is black on the head and back, rusty brown beneath.

FIG. 66.—PRINCE RUDOLPH'S BIRD OF PARADISE (Paradisornis Rudolphi).
This may well be termed the Blue Bird of Paradise, and in its whole coloration is most distinct from all other known species of the family. But as so many of these
strange and beautiful birds have been discovered in the north-western and south-eastern peninsulas of this great island, while a space of about 600 miles long by 300 wide, containing a great snow-topped mountain range, remains wholly unknown, there must be no lack of new and perhaps still more wonderful forms to reward the next generation of explorers and collectors.
In the latter part of the eighteenth century, eight species of these remarkable birds were known, and by the middle of the nineteenth these had been increased to thirteen. But since the occupation of eastern New Guinea by the British and Germans a rapid increase has taken place. In the last edition of my Malay Archipelago in 1890 I was able to enumerate 37 species; and at the present time Mr. Ernst Hartert informs me that 52 species are known, but as he includes two species of Manucodia, which I have omitted from the family, the number of undoubted Paradise birds now known may be safely placed at 50.
Why such wonderful birds should have been developed here and nowhere else is a mystery we shall perhaps never completely solve; but it is probably connected with the absence of the higher types of the smaller arboreal mammalia, many of which are especially destructive to eggs and young birds, together with the protection afforded by luxuriant equatorial forests. The only other country in which similar strange developments of plumage and equally superb colours are found is Equatorial America, where somewhat similar conditions prevail, and where mammalia of a low grade of organization have long predominated. Whatever may be the causes at work, their action has not been restricted to the Paradise birds. Nowhere else in the world are Pigeons and Parrots so numerous and so beautiful as in New Guinea. The great crowned pigeons, the largest of the whole family and rivalling the largest game birds, were first described by Dampier as "a stately land-fowl about the size of the dunghill cock, sky coloured, but with a white blotch and reddish spots about the wings, and a long bunch of feathers on the crown" (figured at the bottom of the plate on p. 424). Many of the fruit-doves are strikingly beautiful, being adorned with vivid
patches of crimson, blue, or yellow, on a pure green ground. Parrots are wonderfully varied, including the great black and the white cockatoos; the lories, varied with crimson and purple, green, yellow, and black; while there are strange little crested green parrots hardly larger than our blue tit—the smallest of the parrot-tribe, as the great black cockatoos are the largest. Kingfishers, too, are remarkably abundant, and include several of the fine racquet-tailed species, with plumage of silvery blue, and with white or crimson breasts. One of these is figured on the plate at p. 424. It is the Tanysiptera galatea of the north-west peninsula. It is bright blue and white, with a coral red bill. Many other groups of birds are also adorned with exceptionally gay colours; and a careful comparison with the birds of other countries shows, that nowhere in the world is there so large a proportion of the whole number of species adorned with brilliant hues. Among insects the same thing occurs, though not in quite so marked a degree; yet the superior beauty of many groups of beetles over the corresponding groups in Borneo is very distinct; and the same is to some extent the case with the butterflies and moths.
Independently of the beauty and singularity, the great number of species of birds inhabiting New Guinea is very remarkable. Considering that there are no resident collectors in the island, and that our knowledge is wholly derived from travellers who have spent a few weeks or months on the extreme northern or southern coasts only, leaving the great mass of the interior wholly unexplored, the number of land-birds already known is surprising.
Mr. Ernst Hartert, the curator of the Hon. L. Walter Rothschild's museum at Tring, has kindly given me the number of birds known to inhabit the Papuan area, that is New Guinea, and those small islands immediately around it and separated from it by a shallow sea either wholly within or only just beyond the 100 fathom line. These are the Aru Islands, Mysol, Waigiou, the islands in Geelvink Bay, and those near the east end of New Guinea. The number of true land-birds now known from these islands
VOL. I. F F
is about 770; and there can be little doubt that within a dozen more years they will reach 800.
If we compare this number with those of other tropical islands on the globe we shall find a remarkable superiority. Next in size is Borneo, long thought to be the largest true island on the globe, and more favourably situated in its proximity to the Asiatic continent; yet, even if we allow for considerable additions during the last ten years, it possesses less than 450 land-birds. Next comes Madagascar, about two-thirds the area of the Papuan islands, yet it possesses barely 200 species of land-birds, although here again its position with regard to Africa seems very advantageous. The whole of the West Indian Islands (excluding Trinidad) forming the Antillean sub-region, has about one-third the area of New Guinea, but being scattered over a larger extent of sea and being in close proximity to two continents from both of which it receives numerous emigrants, would seem to be very advantageously situated, yet it has less than 300 species of land-birds. Then we have the great island-continent of Australia, with an area of close on 3,000,000 square miles, or about nine times that of the Papuan islands, with the advantage of both tropical, sub-tropical and temperate climates, yet possessing less than 500 species of land-birds.
These comparisons render it certain that as regards a rich and magnificent bird-fauna no islands in the world can compare with New Guinea, and it seems doubtful whether any equal area within the tropics contains a larger number of species. When we consider, further, that less than half its area is known, and even that portion very imperfectly, while its most extensive mountain ranges are quite unexplored, it becomes almost certain that New Guinea will ultimately rank as the richest bird-district in the whole world—a very ornithological paradise for the collectors and bird-lovers of the Twentieth Century.
The Papuans.
The peculiar race of mankind inhabiting this great island attracted the attention of the earliest voyagers, and
the country was called New Guinea from the resemblance of its inhabitants to the Negroes of Africa, removed from them by nearly one-third the circumference of the globe. The early writers, however, term the people Papuas or Papuans, a Malay term given to them on account of their woolly hair, so different from the perfectly straight hair of almost all the other Eastern races. The Malay word "papuwah" or "puwah-puwah," means frizzled like wool; and the Malays still call these people "orang papuwah"—woolly-haired men, and the island itself, "tana papuwah"—the land of the woolly-haired.
It is a very remarkable fact that woolly-haired people should be found in two such widely-separated areas, and with very few exceptions, nowhere else in the world. In Africa they occupy the larger portion of the continent, extending over all the tropical and southern regions; while in the East they are found in a group of islands of which New Guinea is the centre, extending westward as far as Flores and eastward to the Fijis. There are also a few outlying groups of woolly-haired people, which are of great importance as indicating that this type once had a wider extension than now. In the Pacific we have the now extinct Tasmanians; and far to the east, in the midst of the brown Polynesians, we find the inhabitants of Penrhyn's Island and Mangaia, in about 158° west longitude, to be of the Melanesian or dark race. In the Philippines there is an aboriginal race of woolly-haired dwarfs—the Aëtas or Negritos; and a similar descriptive term may be applied to the Semangs of the Malay Peninsula, and to the natives of the Andaman Islands in the Bay of Bengal. These various Eastern tribes differ among themselves quite as much as do those of Africa. Both agree, however, in being usually very dark-skinned, and examples may be found in which Negroes and Papuans are in all respects very much alike. But this is exceptional, and there is almost always a characteristic difference which would cause most of the Eastern Negroes to appear out of place on the continent of Africa. The woolly hair, however, combined with the dark skin and almost always with a dolichocephalic or long skull, so markedly distinguishes all these people from the
F F 2
rest of the inhabitants of the globe, that it is impossible not to look upon them as being really related to each other, and as representing an early variation, if not the primitive type of mankind, which once spread widely over all the tropical portions of the eastern hemisphere. Successive incursions of the lighter-coloured, smooth-haired races seem to have exterminated them in many of the areas they once inhabited, while in some widely-scattered spots a few scanty remnants continue to exist. Two important groups, however, remain predominant in regions very far apart, but each well suited to their vigorous development. The Negro of Africa has been made the servant of the more civilized races from the earliest periods of history, and is better known to us than any other uncivilized people; while the Papuan or Melanesian, inhabiting a group of tropical islands on the other side of the globe, still remains a mere shadowy name to the great majority of English readers. We will proceed to point out the chief physical and mental characteristics, habits, and customs, of this interesting race as it exists in New Guinea, with occasional references to such modifications of it as occur in the other islands.
We now possess trustworthy descriptions of the Papuans as they exist at numerous localities scattered all round the extensive island they inhabit: and the substantial agreement of these descriptions renders it pretty certain that all belong to one race, exhibiting, it is true, considerable variations, and occasionally presenting undoubted signs of intermixture with other races, but always showing a decided predominance of true Papuan characteristics. In stature they present a medium between the short Malays and tall Polynesians, the average height varying at different parts of the coast from five feet two to five feet eight inches. Some tribes in the interior are believed to be as dwarfish as the Negritos of the Philippines, while others are nearly equal to the tall Fijians, who are often considerably over six feet high. They are strong and muscular, but rather less finely formed than many of the Malayan and Polynesian tribes. Their colour is usually a chocolate-brown, sometimes almost black, at others almost
as light as some of the Malays. It is, however, by their features that they are best distinguished from most other races of men, and especially by the form and size of the nose. This is always large and long, usually arched as in the Jewish type, and, when well developed, with the extremity so lengthened as to hide the nostrils and overhang the upper lip. This peculiar characteristic is found more or less developed everywhere round the coast of New Guinea, so that almost every traveller speaks of the "Jewish features"—the "aquiline" or "arched" or "very prominent" noses—or makes use of other similar expressions, clearly showing that this is the typical Papuan feature, a fact which is further demonstrated by the unmistakable, though exaggerated, manner in which it is represented in all their images and carvings. The nose is also very thick and coarse, as is the case in almost all savage races, the alæ are very oblique, and the base is much depressed between the eyes, a character which reaches its maximum in the natives of New Caledonia and the New Hebrides, though the nose itself is with them somewhat shorter. The forehead is rather flat and retreating, the mouth large, and the lips full, but not excessively thick; nor is there any marked prognathism. The combination of these peculiarities in various degrees produces faces which are sometimes ugly and savage-looking, while others have so much the character of the Jew or Arab as to be really handsome. Comparing Papuans with typical negroes of Equatorial Africa we find a radical difference in the small flat nose and very prominent jaws of the latter. In the South African races this difference is less pronounced. The Bechuanas and Natal Kaffirs have less prognathism and a straighter, better-formed nose, but this organ is always shorter and less arched than in the Papuan. The Hottentots have often well-formed features and sometimes have a considerable resemblance to the less typical Melanesians. The greatest resemblance, however, is to be found between the Negritos of the Philippines—who have short flat noses and somewhat projecting jaws—and some of the dwarfish tribes of Central Africa.
The Papuan contrasts strongly with Malays and Polynesians in being hairy-bodied and tolerably well bearded, but still more so by the wonderfully luxuriant growth of the hair of the head, which forms a dense mop often projecting six or eight inches from the skull. It is crisp, glossy, and very elastic, and each separate hair naturally curls itself up into a spiral of small diameter. The degree of twist and consequent woolliness of the hair seems to be dependent on its being oval or flattened, instead of cylindrical. In the straight-haired races and in most Europeans the hair has a circular section, which becomes slightly oval where it is naturally curly; but in the Negro and Papuan it is much flattened, and has besides irregular wavy margins, which seem to produce the strong spiral twist. Those who possess a large mop of hair are very proud of it, keeping it continually combed out with a kind of bamboo fork, and using a narrow wooden pillow on which to rest the nape of the neck, so as to preserve the hair from being squeezed out of shape. It was long thought that the hair of these people possessed a peculiar character in growing in separate small tufts scattered uniformly over the scalp; but more accurate examination shows that it grows evenly over the surface of the head, and that the tufted appearance probably arises from the tendency of the spirally twisted hairs to mat together in small curly locks. The hair on the body and limbs, though very short, has the same appearance and a similar structure.
The native of Dutch New Guinea, here figured from a good photograph (Fig. 67), well exhibits the special features of the type—the arched nose with elongated apex, and the mop of frizzly hair. In some individuals the hair is even more developed, but this is rare, while in the majority of the people it is considerably shorter, being always frizzled but of moderate length, as in the group from Southern New Guinea.
The dress of these people is very scanty, the men wearing, the usual T bandage of bark-cloth, but in some cases only a shell, or even going absolutely naked; while the women always wear some kind of girdle, from which is suspended a small apron of bark or a fringe of leaves. As with most
savages, ornament is more attended to than dress, and is more used by the men than by the women. They often pierce the sides of the nose, sticking in them pieces of bone,

FIG. 67.—A NATIVE OF DUTCH NEW GUINEA.
feathers, or tusks of the wild pig. The ears are also pierced, and either shell earrings are worn, or sticks ornamented with feathers are stuck through the lobes. Necklaces of
teeth or shells are common, and heavy rings of white shell or plaited bands of grass, or palm-leaf are worn on the arms. The hair of the men is always carefully attended to. It is combed with a kind of bamboo fork with four or five prongs, and this is usually kept stuck in it both for convenience and ornament. Some tribes cut and trim, or plait the mop of hair into various helmet-like or other fantastic shapes, and all adorn it with combs, sticks, or feather ornaments. Suspended from the neck they often wear a small carved wooden figure with the Papuan features greatly exaggerated. As they freely part with these, they are probably mere ornaments or charms, rather than idols or fetishes. Regular tattooing is unknown, except on the south-eastern peninsula, where there is an infusion of Polynesian blood, but most of the men have raised marks produced artificially. These generally consist of a few short parallel lines on the arms or breast, and are said to be formed by gashes made with a sharp stone or bamboo, and the subsequent application of fire to make the skin swell up and leave a prominent scar. Painting the body is not generally practised, but some kind of stain producing a blue-black tinge has often been observed.
The houses of the New Guinea people are somewhat different in different localities, but the most general type is that found at Dorey Harbour. There is here a considerable village of large houses, built on piles in the water in the usual Malay style, and houses similarly raised on posts (but loftier) are found on the hills some miles inland. Each of these houses is large and accommodates several families, and they are connected by continuous platforms of poles and bamboos, often so uneven and shaky that a European can with difficulty walk on them. A considerable space separates this platform from the shore, with which however it is connected by narrow bridges formed of one or two bamboos, supported on posts, and capable of being easily removed. A larger building has the posts carved into rude forms of men and women, and is supposed to be a temple or council-house. This village is probably very like the pile villages of the stone age, whose remains have been found in the lakes of Switzerland and other countries.
Similar houses are found in the Aru and Ké Islands, in Waigiou, and on the south-west coast; and they are also common on the south-east coast, sometimes standing in the water, sometimes on the beach above high-water mark. These houses are often a hundred feet long, and sometimes much more, and are occupied by ten or twenty families. On the Fly river similar large houses occur, but only raised a foot or two above the ground; while at the mouth of the Utanata river, on the south-west coast, a large low house was found a hundred feet long, and only six feet wide, with nineteen low doors; but this was evidently only a temporary seaside habitation of a tribe which had its permanent dwellings inland.
Finding these large houses, raised on posts or piles and common to many families, to prevail from one end of New Guinea to the other, both on the coast and inland, we are led to conclude that those described by Dr. Miklucho Maclay at Astrolabe Bay, on the north-east coast, are exceptional, and indicate the presence of some foreign element. The houses of the people among whom he lived were not raised on posts, and had very low walls, so that the somewhat arched roofs appeared to rise at once from the ground. They were of small dimensions, and seem to correspond pretty closely to those of the Admiralty Islands, New Britain, and New Ireland; so that this part of the coast of New Guinea has probably been colonized from some of the adjacent islands, a view supported by the fact that these people do not use bows and arrows, so general among all the true Papuans, and by other peculiarities. It is somwhat unfortunate that the only scientific man who has resided alone among these people for more than a year, for the express purpose of studying them exhaustively, should have hit upon a place where the natives are probably not true indigenes but an intruding colony, although perhaps long settled in the country. Dr. Miklucho Maclay will no doubt be quoted as the greatest living authority on the Papuans of New Guinea; and it is therefore very important to call attention to the fact that the people he so carefully studied are not typical of the race, and may not even be Papuans at all in the restricted
sense in which it is usually applied to the main body of the aborigines of New Guinea.
The Papuans, as well as all the tribes of dark, frizzly-haired Melanesians, make pottery for cooking, thus differing from all the brown Polynesian tribes of the Pacific, none of whom are acquainted with this art. Of course the actual seat of manufacture will be dependent on the presence of suitable materials; but those who do not make it themselves obtain it by barter, so that earthenware cooking vessels appear to be in general use all over the island. Cups and spoons are made out of shells or cocoanuts, while wooden bowls of various sizes, wooden mortars for husking maize or rice, wooden stools used as pillows, and many other articles, are cut out and ornamented with great skill. A variety of boxes are made of the split leafstalks of the sago palm, pegged together and covered with pandanus leaves, often neatly plaited and stained of different colours, so as to form elegant patterns. A variety of mats, bags, and cordage are made with the usual skill of savage people; and their canoes are often of large size and beautifully constructed, with high-peaked ends ornamented with carvings, and adorned with plumes of feathers.
The weapons chiefly used are spears of various kinds, wooden swords and clubs, and bows and arrows; the latter being almost universal among the true Papuans and most of the allied frizzly-haired races, while the Polynesians seem never to possess it as an indigenous weapon. It is very singular that neither the Australians, the Polynesians, nor the Malays should be acquainted with this weapon, while in all the great continents it is of unknown antiquity, and is still largely used in America, Asia, and Africa. Peschel, indeed, attempts to show that the Polynesians have only ceased to use it on account of the absence of game in their islands; but mammalia are almost equally scarce in the New Hebrides, where it is in constant use even in the smallest islands; whilst in Australia, where they abound, and where it would be a most useful weapon, it is totally unknown. We must therefore hold that the use of the bow and arrow by the
Papuans is an important ethnological feature, distinguishing them from all the peoples by whom they are immediately surrounded and connecting them, as do their physical peculiarities, with an ancient widespread negroid type.
In their knowledge and practice of agriculture the Papuans show themselves to be far superior to the Australians, and fully the equals of the Polynesian races. They grow cocoa-nuts and bread-fruit, and cultivate various kinds of yam, sweet-potato, bananas, and sugar-cane. Though possessing, for the most part, only stone axes, they clear the forest to make their plantations, which they carefully fence round to keep out the wild pigs. Looking at these clearings, at their houses, their canoes, their implements, weapons, and ornaments often elaborately carved, we must, as Dr. Maclay remarks, be struck with astonishment at the great patience and skill displayed by these savages. Their chief implement, the axe, consists of a hard grey, green, or white stone, made smooth and sharp by long grinding and polishing. A piece of the stem of a tree which has a branch passing off at an angle, something like the figure 7, is hewn off, and upon the branch, which has been cut off short and shaven at the top, the stone is laid horizontally, and bound fast with split rattans or tough bark. Such an instrument requires to be used with great skill, only to be attained by practice, or the stone will be broken without producing any result. These savages can, however, with a stone axe having a cutting edge only two inches broad, fell a tree-trunk of twenty inches diameter, or carve really fine figures on a post or spear. Each adult man possesses one such axe, but in every village there are usually one or two larger two-handed axes, which are about three inches broad. These are considered exceedingly valuable, and are only used for cutting large trees for canoes or other important work. Fragments of flint and shells are used for finishing carved work and cutting the ornamental patterns on bamboo boxes, as well as for making combs, spoons, arrows, and other small articles. For cutting meat and vegetables a kind of chisel of bone and knives
of bamboo are made use of. On the north-west and south-west coasts, where the people have long been in communication with Malay traders, they have iron tools and weapons, and cultivate also maize and a little rice and millet, and have the papaya as an additional fruit and vegetable; and they also grow tobacco, of which they make huge cigars. At Dorey they have learnt to work iron, and make swords and choppers as well as iron points to their arrows and spears.
The daily food of these people consists of some of the vegetables already named, of which they have a pretty constant supply, together with fruits, fish, and occasionally the flesh of the wild pig, the cuscus, or of birds caught in snares or shot with arrows. They also eat shell-fish, lizards, and almost every kind of large insect, especially beetles and their larvæ, which are eaten either raw or cooked. Having no salt, they mix sea-water with that in which they cook their food, and this is so highly esteemed that the people of the hills carry away bamboos full of salt water whenever they visit the coast.
The plantations are usually made at some distance inland for safety, and after the ground is cleared and fenced by the men, the cultivation is left almost wholly to the women, who go every day to weed and bring home some of the produce for the evening's meal. They have throughout the year a succession of fruits and vegetables either wild or cultivated, and are thus never half-starved like the Australians. On the whole the women are well treated and have much liberty, though they are considered as inferiors, and do not take their meals with the men. The children are well attended to, and the fathers seem very fond of their boys, and often take them when very young on their fishing or hunting excursions.
As in the case of most other savages, we have very different and conflicting accounts of the character of the Papuans. Mr. Windsor Earl well remarks, that, whenever civilized man is brought into friendly communication with savages, the disgust which naturally arises from the first glance at a state of society so obnoxious to his sense of propriety, disappears before a closer acquaintance, and
he learns to regard their little delinquencies as he would those of children; while their kindliness of disposition and natural good qualities begin to be recognized. Thus many writers make highly favourable statements respecting the Papuan character and disposition; while those whose communications with them have been of a hostile nature are so impressed with their savage cunning and ferocity, and the wild-beast-like nature of their attacks, that they will not recognize in them any feelings in common with more civilized races.
Many of the early voyagers record nothing but hostility or treacherous murders on the part of the Papuans. Their visits were, however, chiefly on the north-west and south-west coasts, which the Malays have long been accustomed to visit not only for commerce but to capture slaves. This having become a regular trade, some of the more warlike coast tribes, especially those of Onin in McCluer's inlet, have been accustomed to attack the villages of other tribes, and to capture their inhabitants, in order to sell the women and children to the Malays. It is not therefore surprising, that unknown armed visitors to these coasts should be treated as enemies to be resisted and if possible exterminated. Even Europeans have sometimes increased this feeling of enmity through ignorance of native habits and customs. Cocoa-nut trees have been cut down to obtain the fruit, apparently under the impression that they grew wild and were so abundant as to be of little value; whereas every tree is considered as private property, as they supply an important article of food, and are even more valued than the choicest fruit-trees among ourselves. Thus Schouten, in 1616, sent a boat well-armed to bring cocoa-nuts from a grove of trees near the shore, but the natives attacked the Europeans, wounded sixteen of them, and forced them to retire. Commodore Roggewen, in 1722, cut down cocoa-nut trees on the island of Moa on the north coast, which of course brought on an attack. At other times houses have been entered in the absence of their owners, a great offence in the eyes of all savage people and at once stamping the intruder as an enemy.
On the other hand Lieutenant Bruijn Kops, who visited the north-west coast of New Guinea in 1850, gives the following account of the inhabitants of Dorey:—
"Their manners and customs are much less barbarous than might be expected. On the contrary they give evidence of a mild disposition, of an inclination to right and justice, and strong moral principles. Theft is considered by them as a grave offence, and is of very rare occurrence. They have no fastenings to their houses, yet seldom or never is anything stolen. Although they were on board our ship or alongside during whole days, we never missed anything. Yet they are distrustful of strangers until they become acquainted with them, as we experienced. This is probably less, however, a trait of their character than the result of intercourse with strangers who perhaps had frequently tried to cheat them. The men, it is true, came on board from the time of our arrival, but they were very cautious in letting any of the things they brought for sale out of their hands. The women were at first very fearful and fled on all sides when they saw us, leaving behind what they might be carrying; but at length when they found they had no injury to dread from us they became more familiar. Finally, they approached without being invited, but remained timid. The children very soon became accustomed to us, and followed us everywhere.
"Respect for the aged, love for their children, and fidelity to their wives, are traits which reflect honour on their disposition. Chastity is held in high regard, and is a virtue that is seldom transgressed by them. A man can only have one wife, and is bound to her for life. Concubinage is not permitted. Adultery is unknown among them. They are generally very fond of strong drink, but although they go to excess in this, I could not learn that they prepared any fermented liquor, not even sago-weer or tuak (palm wine). Kidnapping is general in these countries, and is followed as a branch of trade, so that there is no dishonour attached to it. The captives are treated well, are changed if there are any of theirs in the enemy's hands, or released on payment of a ransom, as was the case in Europe in the middle ages."
My own experience of the Papuans at Dorey, in 1858, agrees with this account; and as I lived there for four months with only four Malay servants, going daily unarmed into the forests to collect insects, I was completely in their power had they wished to attack me. A remarkable proof of their honesty occurred to me at the island of Waigiou, where a man who had received payment in advance for Red Birds of Paradise, brought back the money, represented by an axe, when after trying for several weeks he had failed to catch any. Another, who had received
payment for six birds, brought me in the fifth two days before I was to leave the island, and immediately started off for the forest to seek another. Of course I never expected to see him again, but when my boat was loaded, and we were just on the point of starting, he came running down to the beach holding up a bird, which he handed to me, saying with evident satisfaction—"Now I owe you nothing." My assistant, Mr. Allen, venturing alone among the mountaineers of the north-west peninsula, found them peaceable and good-natured. Drs. Meyer and Beccari and Signor D'Albertis, penetrating inland beyond Dorey, were never attacked or seriously opposed; and Dr. Miklucho Maclay suddenly appearing at Astrolabe Bay, among people who seem never to have had any communication with Europeans, soon established friendly relations with them, although subject to great trials of temper and courage at the outset.
His experience with them is very instructive. They appeared at first distrustful and suspicious of his intentions, as well they might be. Sometimes they left him quite alone for days together, or kept him prisoner in the little hut he had built for himself, or tried to frighten him by shooting arrows close to his head and neck, and pressing their spears against his teeth till they made him open his mouth. Finding, however, that he bore all these annoyances good-humouredly, and as a medical man took every opportunity of doing them services, they concluded he was a good spirit, a man from the moon, and thenceforth paid him great respect, and allowed him to go about pretty much as he pleased. This reminds us of the experience of the Challenger at Humboldt Bay, where it was decided not to stay, because some of the natives similarly drew their bows at the officers when away in boats. This was no doubt nervous work for the person threatened, but it was only a threat. Savages do not commence a real attack in that theatrical way, and if they had been met with coolness and their threats been laughed at or treated with contempt, such demonstrations would soon have ceased. Of course it requires very exceptional courage and temper, not possessed by one
man in a thousand, to do this; but the fact should be remembered that in many parts of the world such attempts to frighten Europeans have been adopted, but have never resulted in anything serious. Had the Papuans really wanted to rob and murder, they would have enticed the Challenger people on shore, where they would have had them completely at their mercy, whereas those who did go on shore were very civilly treated.
One of the most curious features noticed by Dr. Miklucho Maclay was the apparent absence of trade or barter among the people of Astrolabe Bay. They exchange presents, however, when different tribes visit each other, somewhat as among the New Zealanders, each party giving the other what they have to spare; but no one article seems ever to be exchanged for another of supposed equivalent value. On the whole, the Russian doctor seems to have found these people industrious, good-natured, and tolerably cleanly, living orderly lives, and conforming themselves strictly to the laws and customs which to them determine what is right.
Captain Moresby, Signor D'Albertis, Mr. O. C. Stone, and the missionaries who have explored the south-eastern extremity of New Guinea, have been greatly struck by the apparently quite distinct races they have found there. As far eastward as the head of the Gulf of Papua (on the east side of Torres Straits) the typical Papuans prevail, the natives of the Katow river being described as nearly black, with Jewish noses, and woolly hair, using bows and arrows, and living in houses a hundred feet long elevated on posts—in all respects exactly agreeing with the prevalent type in the western portion of the island. The group of native chiefs shown opposite (Fig. 68) was photographed by Admiral Maclear when surveying on the Alert, and they show how completely the true Papuan type persists in this region, although neither of them has the Papuan mop largely developed. But further east, about Redscar Bay and Port Moresby, and thence to East Cape, the people are lighter in colour, less warlike, and more intelligent, with more regular European features,

FIG. 68.—PAPUANS, SOUTH-WEST NEW GUINEA.
VOL. I. G G
neither making bows nor (except rarely) pottery, and practising true tattooing by punctures—all distinctly Polynesian characteristics. When to this we add that their language contains a large Polynesian element, it is not surprising that these people have been described as a totally distinct race, and have been termed Malays or Malayo-Polynesians. We fortunately possess several independent accounts of these tribes, and are thus able to form a tolerably good idea of their true characters.
Captain Moresby, speaking of the inhabitants of that large portion of the eastern peninsula of New Guinea discovered and surveyed by him, says:—
"This race is distinctly Malayan; but differs from the pure Malay, being smaller in stature, coarser in feature, thicker lipped, with less hair on the face, being in fact almost beardless. The hair on the head is also more frizzled, though this may result from a different dressing. These men have high cheek-bones like the pure Malay; their noses are inclined to be aquiline and sometimes very well formed. Amongst them are met many men with light hair, and what struck us as a peculiarly Jewish cast of features. They rise to a height of from 5 feet 4 inches to 5 feet 8 inches, are sinewy though not muscular, slight, graceful, and cat-like in the pliability of their bodies."1
This description clearly shows that by "Malay" Captain Moresby means "Polynesian," the characters mentioned being in almost every respect directly the opposite of those of the true Malays, as indicated by the words and phrases here placed in italics. And even as compared with the typical brown Polynesian, the frizzled hair, aquiline noses, and Jewish cast of features, are all Papuan characteristics.
Mr. Octavius C. Stone described the Motu tribe who inhabit the coast districts about Redscar Bay and Port Moresby as somewhat shorter than the Papuans to the westward, and of a colour varying from light brown to chocolate. The hair varies from nearly straight to woolly, often being frizzled out like that of the typical Papuan. The hair on the face is artificially eradicated, and they are thus made to appear beardless. The nose is aquiline and thick, and in a small percentage of the men the Jewish
1 Journal of the Royal Geographical Society, vol. xlv. p. 163.
type of features appears. The adjacent tribes differ somewhat. The Koiari, Ilema, and Maiva are generally darker in colour; while the Kirapuno are lighter. These last live near Hood Point, and are the handsomest people in New Guinea. Their hair is of a rich auburn, often golden in the children, growing in curls or ringlets. It is this tribe that keep their villages in such excellent order, with well-kept gardens in which they even cultivate flowers. Mr. Lawes says: "we were all amazed at the cleanliness, order, and industry, which everywhere declared themselves in this model New Guinea village. The men are physically very fine and the women good-looking. One of the belles of the place had no less than fifty-four tortoise-shell earrings in her two ears, and her nose pierced too."1
Speaking of all these tribes as forming essentially one race, Mr. Stone says, that they are a merry laughter-loving people, fond of talking, and loving a joke, hot of temper, and quick to resent a supposed injury—all of which are Polynesian or Papuan as opposed to Malayan characteristics. They are clean in their habits, and particularly so in their eating. When allowed liberties they do not fail to take advantage; and, at Port Moresby in particular, they are accomplished thieves, inveterate liars, confirmed beggars, and ungenerous to a degree, so that, even if starving, they would give you nothing without an equivalent. This condemnation, however, does not apply to the interior tribes who have not yet been demoralized by European visitors. Both sexes are vain of their outward appearance, oiling their bodies, and adorning themselves with shells, feather and bone ornaments; and on all festive occasions each tries to outvie the other in his or her toilet. Their dress is like that of the Papuans, a T bandage for the men, a fringe of leaves for the women, but the latter are more carefully made than among the more savage tribes. They practise true tattooing, the women especially being often highly ornamented with complex patterns on the body and limbs, and occasionally on the face also, but wanting the elegant curves and graceful designs which characterize
1 Journal kept by Mr. Lawes, Times, November 27, 1876.
G G 2
Polynesian tattooing.1 Their weapons are spears, shields, stone clubs, and hatchets, one tribe only—the Ilema—making bows and arrows. In like manner the Motu tribe only make pottery, which the other tribes obtain from them by barter. They use drilling machines with a spindle wheel and cord, like the Polynesians. The houses, whether on shore or inland, are raised on piles, but are small as compared with those of the Papuans, each accommodating one or two families only.
Intellectually these people are considerably advanced. They can reckon up to a million. They use the outstretched arms as a unit to measure by. They divide the year into thirteen months, duly named, and reckoned from the new moons. The four winds and many of the stars have names, as well as every tree, shrub, flower, and even each well-marked grass and fern. They prefer fair to dark people, and are thus disposed to like and admire the white races. The children are very merry, and have many toys and games. The Rev. W. Turner tells us that they make small windmills of cocoa-nut leaves, and are well versed in the mysteries of cat's cradle; while spinning a button or round piece of shell on a cord, and keeping a bladder in the air by patting it with the hands, are favourite games. They also amuse themselves with miniature spears and bows and arrows, catching fish, which they cook for themselves on the shore. They are left to do what they like, and know nothing of the tasks of school, the difficulty of keeping their clothes clean, or the misery of being washed—troubles that vex the lives of almost all civilised children. According to Mr. Turner, the villages of the Motu are by no means clean, all manner of filth being left about unheeded; and as this agrees with most other descriptions, we must conclude that the model village already referred to is quite exceptional in its cleanliness and order.
Mr. Turner thinks the Motu are colonists from some other land, while he considers the Koiari of the interior to be "evidently the aborigines of this part of New
1 See figures illsutrating the Rev. W. Turner's article on "The Ethnology of the Motu," in the Journal of the Anthropological Institute, 1878, p. 480.
Guinea." Mr. Stone, on the other hand, classes them together as slightly differing tribes of the same race, the one being a little more advanced than the other; and he considers the whole eastern peninsula of New Guinea to be peopled by a race of Polynesian blood, who, in some far distant time, found their way to the coast, intermingled with the native Papuan tribes, and gradually drove them westward. There has thus resulted a number of separate tribes, showing various degrees of intermixture, the Polynesian blood predominating on the coast, the Papuan in the interior; one small tribe alone, the Kirapuno, being more distinctly Polynesian. How complete is the intermixture, and how difficult it is to determine the limits of the two races, is shown by the opinion of Mr. S. M'Farlane, who says, that though he at first thought the people of Katow River and those of Redscar Bay to be quite distinct, the former Papuan and the latter Malayan (or more properly Polynesian), yet, after five years' acquaintance with them, he believes them to be of the same race; while he considers the tribes of the interior to be distinct, and to be true Papuans. The coast people he thinks to be the result of an intermixture of Malays, Polynesians, Arabs, Chinese, and Papuans.
Dr. Comrie (of the surveying ship Basilisk) believes that all the tribes on the north-east coast, from East Cape to Astrolabe Bay, are Papuans; but his description of them shows that they have a slight infusion of Polynesian blood, and many Polynesian customs. One thing is very clear, that neither in physical nor mental characteristics do these people show any resemblance whatever to Malays, who are a very different race from the Polynesian. The graceful figures, the woolly or curly hair, the arched noses, the use of tattooing, the ignorance of pottery-making, the gay and laughter-loving disposition, the talkativeness of the women, the lying, thievishness, and beggary, widely separate them from the Malay; while all these peculiarities support the view of their being a race formed by a mixture of Polynesian men with Papuan or Melanesian women, the former having perhaps arrived in successive waves of immigration, thus causing the coast tribes, and those
nearest the eastern end of the island, to be more distinctly Polynesian in character than those inland and towards the west.
Returning now to the dark Papuan tribes of the remainder of New Guinea, we find that here also there is some difference of opinion. Owing to the coast tribes being usually at war with those of the interior, these latter have been described by them as a different race, and have been called by the Dutch and other writers Alfuros1 or Harafuras, a term applied to any wild people living in the interior of a country, as opposed to the coast tribes. This has led many writers to class the natives of New Guinea into Papuans and Harafuras, terms which are still sometimes used, but which are quite erroneous as implying any physical difference or any distinction of race. Dr. Meyer, who has seen much of the people of the north-west coast, considers that there is no difference of the slightest importance between the coast and inland tribes, but such as occur in every race. Dr. Miklucho Maclay concludes that the Papuan stock consists of numerous varieties, with no sharp lines of demarcation. Dr. Beccari, however, differs somewhat from the preceding writers; and as he explored a great range of country, and made repeated visits to the western half of New Guinea, his opinion is entitled to great weight. He thinks there are three distinct types of Papuans. One is dwarfish, with short woolly hair, skin almost or quite black, nose much depressed, forehead extremely narrow and slanting, and with a brachycephalous cranium; these he terms Oriental Negroes or Primitive Papuans. They do not now exist as a race, but are scattered among the interior tribes, and their description accords very closely with that of the Negritos of the Philippines and the Semangs of the Malay Peninsula. The next are the typical Papuans, who are most widely spread, and present most of the characteristic features we have already described. The last are the Mafu or Mafor Papuans who inhabit Dorey and the shores and islands of Geelvink
1 The term is derived from the Portuguese "fora," out or outside; Alfores being applied to tribes out of or beyond the settlement on the coast (Windsor Earl's Papuans, p. 62).
Bay, and are probably scattered all round the western coasts. They form the highest type, with fine Jewish or European features, a better intellect, and a somewhat more advanced civilization. These people divide the year into lunar months, each with a proper name, and have names for the four cardinal points, for many stars, and for entire constellations. Dr. Beccari believes them to be the result of an intermixture (at a remote epoch) of Hindoo or Caucasian blood with the indigenes of the island, and he even traces a connection between their rude mythology and that of the Hindoos.
A curious point of physiological detail may here be noticed as lending some support to this theory. Almost all observers have remarked, that the fully developed Papuan mop of hair is not a general feature in any of the tribes, but occurs sporadically over a wide area, is highly valued by its possessors, and from its extreme conspicuousness is always noticed by travellers. No other race of people in the world possesses this character at all; but, strange to say, it appears very fully developed among the Cafusos of Brazil. These are a mixed race, the produce of Negro and Indian parent, and their enormous wigs of frizzly hair have been described by Spix and Martius, and are known to most South American travellers. Still more interesting is the appearance of a similar peculiarity among the Arab tribes of Taku in Eastern Africa, where mixtures of Negro and Arab blood are very common.1 It is well known that hybrid and mongrel characters are liable to great variation and are very uncertain in their appearance or degree of development. If, therefore, the higher type of Papuans are the result of a remote intermixture of Hindoos or Arabs with the indigenous Papuans, we can account both for the appearance of the great mop of frizzly hair and for its extremely unequal development; and it is not improbable that the Jewish and greatly elongated nose may have a similar origin.
Outside New Guinea and its closely surrounding islands the same race is found in the Solomon Islands, the New Hebrides, and the Fiji Islands. The native of Alu figured
1 Waitz's Anthropology. English translation, vol. i., p. 175.

FIG. 69,—PAPUAN OF NEW HEBRIDES.
by Mr. Woodford in his Naturalist among the Head Hunters is an almost typical Papuan. The portrait opposite of a man of the New Hebrides (Fig. 69) might be taken for a handsome Jew, were it not for his frizzly hair; and a native

FIG. 70.—NATIVE OF API, NEW HEBRIDES.
of Api in the same group of islands (Fig. 70) has quite a European countenance. The Fijians are equally handsome, with typical Papuan features and hair, and they are often over six feet high. Westward from New Guinea the same

FIG. 71—NATIVES OF TIMOR.
type in a somewhat modified form is found in Timor and Flores, the group of Timor men here shown (Fig. 71) indicating a divergence, in the less frizzly hair a rather.
lighter coloured skin and less pronounced features, but yet on the whole decidedly Papuan. In the islands of Ceram and Gilolo also the type is more Papuan than Malayan, though the light brown colour indicates, perhaps, some intermixture with people of Malay or even of Caucasian race.
If we now take account of all the evidence yet obtained, we seem justified in concluding that the great mass of the inhabitants of New Guinea form one well-marked race—the Papuan—varying within comparatively narrow limits, and everywhere presenting distinctive features which separate it from all other races of mankind. The only important deviation from the type occurs in the southeastern peninsula, where a considerable Polynesian immigration has undoubtedly taken place, and greatly modified the character of the population. At other points immigrants from some of the surrounding islands may have formed small settlements, but it is a mistake to suppose that there are any Malay colonies on the south-west coast, though a few of the natives may have adopted the Malay dress and some of the outward forms of Mahometanism.
If we look over the globe for the nearest allies of the Papuans, we find them undoubtedly in Equatorial and Southern Africa, where alone there is an extensive and varied race of dark-coloured, frizzly-haired people. The connecting links are found in the dwarfish, woolly-haired tribes of the Philippines, the Malay Peninsula, and the Andaman Islands; and, taking these altogether, we may well suppose them to represent one of the earliest, but not, probably, the most primitive type of man.
We must not fail to take note of the fact that the two great woolly-haired races are almost entirely confined within the tropics, and both attain their highest development near the equator. It is here that we should expect the primitive man to have appeared, and here we still find what may be one of his earliest divergent races thriving best. We may, perhaps, even look on the diverse types of the three great races as in part due to changes of constitution adapting them to cooler or warmer climates and
changed conditions;—first, the Australians and the hill tribes of Central India, who once perhaps spread far over the northern hemisphere, but have been displaced by the Mongoloid type which flourishes at this day from the equator to the pole. These, again, have been ousted from some of the fairest regions of the temperate zone by the Indo-Europeans, who seem only to have attained their full development and highest vigour when exposed to the cold winds and variable climate of the temperate regions.
If this view is correct, and the Papuans really form one of the earliest branches of the primitive type of man which still exists on the globe, we shall continue to look upon them with ever-increasing interest, and shall welcome every fact relating to them as important additions to the history of our race. The further exploration of their beautiful and luxuriant island will, it is to be hoped, be vigorously pursued, not only to obtain the mineral, vegetable, and animal treasures that still lie hid in its great mountain ranges, but also to search for the remains of primæval man in caves or alluvial deposits, and thus throw light on the many interesting problems suggested by the physical peculiarities and insular position of the Papuan race.
CHAPTER XXI
AFFINITIES AND ORIGIN OF THE AUSTRALIAN AND POLYNESIAN RACES1
THE aborigines of Australia differ remarkably from those of all the surrounding countries, while they agree so closely among themselves in every part of the continent that they evidently form a single race. To summarize their main characteristics, they may be described as men of medium stature, muscular, but with slender arms and legs, rather large heads, with broad foreheads and overhanging brows, the nose thick and very broad at the nostrils, as in most of the lower races, the mouth large and lips thick, but less so than in many negro tribes. In colour they are a deep copper or chocolate, never sooty black as in the negro; the hair is long, glossy black or very deep auburn, usually wavy or curled, and very abundant, and the face is adorned with a luxuriant growth of moustache, beard, and whiskers, usually with an auburn tinge. These characters in their combination give the face, as a whole, a familiar appearance, resembling that of the coarser and more sensual type of Western Europeans, while they are thereby totally removed from any of the beardless Malayan and Polynesian tribes, or the woolly or frizzly-haired Papuans. (The reproduction of the photographs of the man from North Australia, and the woman from
1 The first portion of this article (to p. 473) forms the concluding section of my chapter on "The Aborigines of Australia" in Australasia, vol. i., and is reprinted here by the kind permission of Mr. Edward Stanford.

FIG. 72.—TUNGAGORA, WORKII TRIBE, NORTHERN TERRITORY.
Queensland, kindly lent me by the Council of the Anthropological Institute, show these characteristics, especially

FIG. 73.
Australian, Queensland. (Collection Anthropologique du Prince Roland Bonaparte.
that of the wavy, glossy hair, equally removed from the frizzly mop of the Papuan as from the straight, coarse hair of the Malay.) They have sometimes a considerable
growth of hair on the body, especially on the chest and back, while the infants are much lighter in colourthan the adults, and are clothed with a soft fur on the neck and back.
If we turn to habits and customs for some light as to the probable derivation of the Australians, we find equally clear proofs that we must go far beyond the limits of all the immediately surrounding peoples. While Malays, Papuans, and Polynesians all cultivate the ground and all build good permanent houses, the Australians never do one or the other. The pottery of the Malays and Papuans, the bows and arrows of the Papuans and other Melanesians, and the well-made and often elaborate canoes of all these races are equally unknown to the Australians, who must thus be classed among those savage peoples which have made the very smallest advance towards material civilization.
Supposed African Affinities.
In Mr. Curr's important work on "The Australian Race," an attempt is made to show that there is a great resemblance in language and in many of their customs, between the native races of Australia and Africa. By comparing words chosen among the numerous languages of Australia with words having the same meaning in some of the still more numerous African languages and dialects, a considerable number of curious resemblances are found. But it is not improbable that an equal number of resemblances could be found if the languages of any other equally extensive portion of the world were taken for comparison, because many words in all languages are either onomatopœic—that is, sound-imitations, or, what may be called functional. Thus, words for the lips, mouth, or head often contain labial sounds; those for the teeth or for things that resemble teeth, contain dentals—that is, sounds which are only produced when the tongue touches the teeth, as d and t; while the nose, and things supposed to resemble it, are named by words containing such nasal sounds as n, ng, or ñy; hence names of these parts of the body and words derived from them will often
resemble each other in the most remote parts of the world, and in languages which in most respects are quite unlike each other. So, the words for father and mother—the infantile ba, pa, ma—occur in every part of the world, though sometimes transposed in their meaning. Similar resemblances to the Australian have been found by other writers among the languages of the hill tribes of India, and with these peoples there is much more agreement in physical characters.
Mr. Curr also adduces a number of curious customs which are common to Australia and Africa, such as the great importance attached to sorcery; the forming raised scars on the body; the habit of knocking out the front teeth; the practice of a form of circumcision; the prohibition of marriage within certain class-divisions; the forms of burial, and a few others. But some of these customs are very widespread among savages, and unless it can be shown that a considerable number of them are strictly limited to Australia and Africa they can afford no proof of a common origin of the two races. More interesting is the fact that the peculiar Australian weapon, the boomerang, finds its nearest representative among the Abyssinians and the ancient Egyptians, though it is doubtful whether it had with these people the same peculiarities of returning and erratic flight. It is possible that the weapon was more widely spread in early times, but was gradually superseded by the bow and other superior weapons; but it can hardly be held to prove any identity of race.
Resemblance of Australians to the Ainos.
If we look broadly, and without prejudice or prepossession at the physical features of the Australians, it is evident that they cannot be classed either with the Negroid or the Mongolian types of man. There remains only the Caucasian, and, though of a very low type, in all essential features they agree with these; and if we look abroad for other isolated fragments of the same race, we find one of them in the Ainos, or hairy people, of Japan. These curious people agree in many respects with the best types
VOL. I. H H
of Australians, but they are somewhat more hairy, of a much lighter colour, and with more refined features. Some

FIG. 74.—ARUNTA TRIBE. OLD MAN, VERY HAIRY.
of the old men resemble handsome European peasants. They are also much more advanced in material civilization than the Australians.

FIG. 75.—HAIRY AINO, URAP.
(The two photographs, of a man of the Arunta tribe in Central Australia, and of a typical hairy Aino, show a
H H 2
remarkable similarity in this respect, and though the Aino is of a more refined type, there is nothing to indicate any radical diversity of race between them.)
Other fragments of the same great primitive type are to be found in the Khmers of Cambodia, as well as in some of the wild tribes of the same country and of Western China, who have long been recognized as of Caucasian type. The language of the Khmers is said to have some affinities with that of the Mahoris, who are, as we have stated in the last chapter, now generally recognized as of Caucasian affinity, as are some of the tribes of Micronesia.1
The Australians are low Caucasians.
Of all these widely scattered Caucasian fragments we must look upon the Australians as the lowest and most primitive. Their antiquity is, in all probability, very great, since they must have entered their present country at a time when their ancestors had not acquired the arts of making pottery or of cultivating the soil, of domesticating animals, of constructing houses, or of fabricating the bow and arrow. They thus afford us an example of one of the most primitive types of humanity yet discovered; and if, as I believe, they really belong to our own stock, we are led to the conclusion that the differentiation of the three great races of mankind took place at so remote an epoch that we cannot expect to find any of the earliest connecting links, while the amount of intermixture that has taken place wherever two of these races come into contact renders their actual delimitation still more difficult.
The uniformity of the low stage of material civilization in the Australians, though spread over so wide an area differing considerably in climate and vegetation, and in some parts very fertile, is the more remarkable, because
1 In Mr. Richard Semon's work on Australia (p. 237) he compares the Australians to the Veddahs of Ceylon and some of the Dravidians of Central India, and he considers that the Caucasians have descended from these races. He adds:—"Several observers have already stated that the physical features of the Australians, in spite of their so-called ugliness and coarseness, may be decidedly compared to the lower Caucasian type."
I published this view in 1893, and have not myself seen it alluded to at any earlier date.
they must again and again, have been brought into contact with more advanced races. There are clear signs of intermixture in the north both with Malays and Papuans, yet this has had little or no effect on the customs of the people. But some higher race has evidently at one time formed a settlement on the north-west coast, as indicated by the very remarkable cave paintings and sculptures discovered by the late Sir George Grey in his earliest exploration of that country. They were found in the valley of the Glenelg River in North West Australia, about sixty miles inland and twenty miles south of Prince Regent's River, in a very rugged tract of country. The paintings consist of representations of human heads and bodies, apparently of females clothed to the armpits, but the faces are without any indication of mouths. The heads are surrounded with a broad head-dress or halo, and one of the figures wears a necklace. They are executed in red on a white ground, the clothing being marked with a red pattern, and the broad hat or halo in some of the figures is coloured blue, red and yellow. These designs are nearly life-size, and the largest is on the sloping roof of a cave thus appearing to look down on a visitor. There are also some drawings of kangaroos, more finished than any of the work of modern Australians. On the roof of another cave there was a full-length figure, ten feet high, dressed in a loose red garment from neck to ankles, the hands and feet being well executed and the latter apparently, covered with shoes. The white face is mask-like, showing the eyes only and around it are circular concentric bandages, the inner one yellow, the outer red, looking something like a broad cap and outer bonnet. On the upper part of this are five letters or characters, having an oriental aspect. Although poorly executed, these figures have a refined character, as utterly out of place among the Australian natives as would be any modern work of art. Very near one of these caves there was found on a large vertical sandstone rock a well-executed human head, hollowed out to about an inch and a half deep in the centre, the whole head being two feet in length and sixteen inches broad. The singularity of it is that it is perfectly European in type, both in form of
head and features. The only other paintings which appear to have a similar character are those discovered by Captain Flinders in Chasm Island on the North East Coast, and which have been preserved in a sketch by W. Westall, A.R.A., who accompanied Flinders as artist. These form a long procession of human figures of a pale red colour, rudely executed, but all apparently clothed in long robes. Near the head of the procession is a much taller male figure, with arms outstretched and holding a stick, and towards the middle is an equally tall female figure. There are also a turtle and a sword-fish, pretty well drawn.
Whoever were the people who executed these singular paintings, they were, probably, also the makers of the two large square mounds found by Sir George Grey in the same district. These were formed of loose stones, but were perfect parallelograms in outline, and were placed due east and west. From the drawing given of them they must have been heaped up with great care, since they are finished to a sharp ridge, with triangular ends just like the roof of a building. Both were exactly the same length, twenty-two and a quarter feet, but they differed somewhat in width and height. One was opened, but nothing was found inside but a quantity of fine mould.
In the same district Sir George Grey noticed among the dark-coloured natives a few individuals who were very much lighter in colour—he says "almost white;" and he thinks that these lighter people exercised authority; and he also describes the native houses or shelters as being better constructed here than in other parts of Australia.1
A good deal further south, on the Upper Gascoyne River, Mr. Giles met with some natives who were exceptionally good-looking. He says: "Some of these girls and boys had faces, in olive hue, like the ideal representation of angels; how such beauty could exist among so poor a grade of the human race it is difficult to understand, but there it was."
These various indications compel us to believe that one or more colonies have formerly existed on the north-west
1 See Journals in North-West and West Australia, by Lieutenant George Grey (afterwards Sir George Grey) 1841.
coast, either of shipwrecked Europeans or of some of the higher and more civilized Malays, with possibly Arabs or Chinese among them, and that, after maintaining themselves for some time and leaving behind them the curious paintings, carvings, and symmetrical stone heaps here described, they have either been able to escape or have been exterminated by the natives, leaving however behind them some mixed descendants to puzzle the traveller by their superior appearance. The remarkable costume of the persons represented in the wall-paintings should give a clue to the designers, and there is, so far as I know, only one not very remote locality where a similar costume is in use—the islands of Siau and Sanguir north of Celebes—but as these people were converted to Christianity by the Portuguese, those I saw at Menado perhaps wore an imitation of the cassock of a Romish priest. It is just possible that some Chinese Christians, converts of the early Jesuit missionaries, may have been wrecked on this coast, and that the figures may be due to their recollections of the images or pictures of saints with haloes round their heads. But whoever the people were who executed these drawings it is quite certain they were not Australians.
The conclusion here reached, that the Australians, though usually classed as one of the lowest forms of mankind now surviving, are really one of the primitive types of the great Caucasian race to which we ourselves belong, will appear to many of my readers to be improbable, or even absurd. But I venture to think that it, nevertheless, most nearly accords with, all the facts of the case; and since it has been admitted that some of the darkest Hindoos are nearly allied to Europeans, there is less improbability in the existence of some more archaic and less developed forms of the same fundamental type. It also accords with all we are now learning of the vast antiquity of the human race, since if all the peoples now living upon the earth can be classed in one or other of the three great divisions of mankind—Negroid, Mongolian, and Caucasian—or as probable mixtures of them, we are impressed with the conviction that we must go back to periods as com-
pared with which the earliest historical dates are but as yesterday, in order to arrive at an epoch when the common ancestors of these three well-marked types alone existed. Even then we shall have made no perceptible approach to the missing-link—the common ancestor of man and the higher quadrumana.
It has been estimated that when Australia was first colonized by Europeans, the native population could not much have exceeded 150,000. They have since greatly diminished, owing to the occupation and settlement of all the more fertile parts of the country, as well as from the diseases and vices introduced among them by the convicts and lower class of settlers. Notwithstanding all these causes of depopulation, great numbers still roam over the interior, and it is believed that they now amount to from 30,000 to 40,000; and as so much of the country they inhabit is not of a nature to invite occupation by the white race, it seems not improbable that the degraded Australians may continue to exist long after the much higher Mahoris of New Zealand and Eastern Polynesia have disappeared as completely as have the lower Tasmanians.
Mental and Moral Characteristics.
The intellectual status of the Australian native has been greatly depreciated by many writers, but more particularly by those who are most ignorant of him in his native wilds, or who judge his mind solely by a reference to his low state of material advancement. Those who have lived much among the natives, however, often express a different opinion. Sir Thomas Mitchell did not think them by any means the lowest in the scale of humanity. Mr. Eyre found them to be frank, open, and confiding, and easy to make friends with; and he declares that they have been greatly misrepresented and traduced, and that much of their assumed treachery and bloodthirstiness is the result of the cruelty and ill-treatment of the settlers. They often show great affection for their male offspring, giving up to them the choicest food, carrying them when they are weary, and lamenting their death for months, or even for years.
Mr. Curr, who for more than forty years was in constant communication with them, and having been for many years "Protector of the Aborigines" in Victoria had excellent opportunities for observation, thus describes them: "The black, especially in his wild state, is quicker in the action of his mind, more observant, and more self-reliant than the English peasant. … Socially the black is polite, gay, fond of laughter, and has much bonhomie in his composition. … Touching the moral feelings of the blacks, observation has convinced me that they are not without them, though they are much blunted by constant repression, and that they discriminate between right and wrong."
It has been observed that the natives are very strict in obeying their laws and customs, even when alone among Europeans, where the offence would be unnoticed, and even under great temptation. The horror of marrying a woman within the prohibited degrees of relationship, the extreme grief they manifest at the death of children or relatives, and sometimes even for white men, as illustrated by the native boy who was the sole companion of the unfortunate Kennedy when he was murdered, are sufficient to indicate that they possess affections and a sense of right and wrong not very different from our own.
(The photograph on the next page (Fig. 76) of a family group at their ordinary occupations, with the two children watching their father at work, may serve to illustrate the foregoing observations as to their character and family affections.)
Australians and other Eastern Caucasians.
The preceding account was written about the year 1892, at which time I had seen no good photographic portraits of the best types of Australians, the majority of travellers appearing to select the most hideous old people they could meet with as illustrations of the race. Having now had the opportunity of examining the fine series of photographs obtained by Professor Spencer for his work on Central Australia, and others from the collection of the

FIG. 76.—FAMILY GROUP. ARUNTA TRIBE, CENTRAL AUSTRALIA.
Anthropological Institute, and of comparing these with the portraits in Mr. Romyn Hitchcock's monograph on the Ainos of Yezzo, Japan, and of Todas from the Nilgherries in Captain Marshall's work on those people, and also in Professor Keane's "Man, Past and Present," I have become more satisfied of the correctness of the view then put forth, I believe for the first time. Professor Keane considers that there are several fragments of primitive Caucasian peoples to be still found in South-Eastern Asia. Such are the Khayems, near the sources of the Irawaddi, who are described as having oval faces, pointed chins, and aquiline noses, and as being sometimes so light in colour as almost to pass for Europeans. Among the Shans of Upper Burma there are also found families or small tribes having distinct Caucasic affinities. But it is in Cambodia where the most striking examples are found in the Khmers and several other tribes, which are believed to be the remnants of an ancient Caucasic race of Hindu or S. Indian affinities which once occupied much of South-Eastern Asia, and were the builders of the grand cities and temples whose ruins exist in the interior forests. So little is known in this country of these interesting peoples, and the idea is so prevalent that all Eastern Asia, from Burma to China and Japan, to Java and the Moluccas on the south-east and to the extremity of Manchuria and Siberia on the north-east, is wholly peopled by men of Mongolian race, that some further account of these Caucasian tribes will be of interest, especially as they help us to a solution of the problem of the origin of the two great races—Polynesians or Mahoris and Australians, which wTe are here discussing.
Although ancient temples of Eastern Asia may seem out of place in a chapter dealing with the origin of the Australians, yet, from the point of view of the theory here suggested, they will be found to have a direct bearing upon the problem. And as the works containing descriptions and plates of these wonderful ruins are unknown to most readers, no further apology is needed for their introduction here.
The Ancient Civilization of the Khmers.
Although a Chinese traveller described this civilization, in all its grandeur only a short time before its

FIG. 77.—PART OF ANCIENT CAMBODIAN TEMPLE AT ANKHOR-THOM.
destruction about six hundred years ago, the wonderful ruins of its cities and temples were first made known to Europe by Mr. Henri Mouhot, a French traveller and natur-
alist, who visited Cambodia in 1858 and died there in 1861. His plans, sketches, and descriptions of them were, however, published in France, and an English translation in 1864. Later on Mr. J. Thomson photographed some of the ruins, and the more important of these were engraved in Ferguson's "History of Architecture," one of the fragments of the great temple in the deserted city of Ankhor-Thom being here reproduced. (Fig. 77) It was not, however, till Garnier and Delaporte made their careful explorations, the results of which were published in Paris in 1873 and 1880, that we obtained any adequate idea of the vast extent, great artistic talent, and wonderful mechanical skill of this interesting people, by means of the accurate plans, photographs and restorations of these ancient monuments given in their works.
Delaporte, in his Voyage au Cambodge informs us that the ruins of Ankhor-Thom, the ancient capital of the Khmers, show that it occupied a space of about eight square miles, and was surrounded by a wall thirty feet high, backed by a thick earthen rampart. Outside the wall there was a deep and wide ditch crossed by several bridges whose parapets were supported by five hundred statues of giants. A special gate, profusely ornamented, led to the great temple of Baioñ, which was so buried in tropical vegetation at the time of Delaporte's visit that he had to employ over sixty workmen for two weeks in clearing away the undergrowth, etc., so as to enable him to draw plans and obtain clear views of the whole structure.
He describes this temple, the centre of the Brahminical religion of ancient Cambodia, as being about 340 feet square, but surrounded by a highly ornamented double cloister, upwards of 500 feet square. The central tower of the temple is 165 feet above the platform of the outer court and its surrounding cloisters, which are themselves considerably elevated, so that the temple was about 200 feet above the adjacent parts of the city.
Captain Delaporte, who was a student of architecture and excellent draughtsman, thus sums up the main impression produced by it. Speaking of the hundreds of
gigantic faces of Brahma which form the unique and characteristic feature of the temple, he says:—
"There is upon these faces—which are somewhat strange but yet regular in their characteristics—an air of force and half-smiling serenity which has a very original air of nobility. Whether near or far off they take their place naturally in the curves of the whole design, and combine in an architectural whole which, notwithstanding the abundance of decorative accessories, still remains true in outline, harmonious in proportion, and grand in general effect."
From the elevation of the temple drawn to scale these faces appear to be about twelve feet high, and as there are fifty-one towers thus ornamented, the centre one having ten, the others four each, there must be in all 210 of these huge portraits.
The great temple of Baioñ is the best designed, the most complex in its forms, and the most original of all the sacred buildings of ancient Cambodia. It at once astonishes and charms, and affords a striking example of the exceptional talent of the bold architects of Khmer. The imposing effect of the whole—as well shown in the view opposite, greatly reduced from an engraving in the atlas to Garnier's work—depends upon the skilful grouping of the fifty-one massive yet highly enriched towers and turrets of a tapering conical outline; while the profusely decorated galleries and cloisters connecting and surrounding these, now mostly in ruins, contained on the inner surfaces of their walls nearly a mile in length, about eleven thousand figures of men and animals. The temple was also decorated with a great number of statues of gold, silver, copper, stone or wood, the latter usually gilt.
The great palace in the same city was equally large, perhaps as beautiful, but in a somewhat lighter and less highly decorated style. These are both very ancient. According to old native history the temple was built 250 B.C.: not very long after the conquest of the country from India; the palace about three hundred years later. Their marvellous preservation is due partly to their mode of construction, partly to the fact that they were only abandoned about three hundred years ago, so

FIG. 78.—TEMPLE OF BAIOÑ, CAMBODIA.
that the destroying effect of tropical vegetation has only been acting on them for that short period.
The mode of construction of these buildings is thus described by Ferguson, whose long-continued personal study of Indian architecture constitutes him an authority of the first rank. He says:—
"Nothing can exceed the skill and ingenuity with which the stones of the roofs are joggled and fitted into one another, unless it is the skill with which the joints of their plain walls are so polished and so evenly laid without cement of any kind. It is difficult to detect their joints even in a sun-picture which generally reveals flaws not to be detected by the eye. Except in the works of the old pyramid-building Egyptians, I know of nothing to compare with it. When we put all these things together, it is difficult to decide whether we ought most to admire the mechanical skill which the Cambodian architects displayed in construction or the largeness of conception and artistic merit which pervades every part of their designs."
The stone used in the construction of these temples and palaces is a fine grained sandstone, so hard and compact as to take a polish; and as no lime or cement was used, and the joints are so wonderfully close, there was little opportunity for seeds to vegetate except in the hollows of the ornamental carving of the numerous turrets, where the view shows a considerable growth of dwarf vegetation. Some of the surrounding cloisters and perhaps other parts of the building seem to have been wilfully destroyed, perhaps at the time of the Siamese conquest, and in such broken portion forest trees have grown, and their roots penetrating among the foundations, have so loosened them that some of the towers have been destroyed. Delaporte tells us that one turret fell while he was encamped there, and he thinks that unless protected the whole building will soon be irretrievably ruined. It is to be hoped, however, that the French government will see that this magnificent ruin is carefully preserved for future generations. Great numbers of other ruins in about fifty distinct sites, are scattered over a district extending for 400 miles in a north and south direction and 200 miles east and west, occupying nearly all of Cambodia with portions of Siam and Cochin China.
Among these are numerous low pyramids, often surmounted with lofty towers, and a great variety of other buildings now mostly in ruins. Other temples are of much later date, and exhibit an architectural style which seems a combination of Cambodian and Roman.
The people who built these magnificent temples and cities, and also made great lines of roads equal to those of the Romans, with bridges which, in some cases, are still in perfect preservation, are the Khmers, who still live in the country and preserve their nationality, though surrounded by Siamese, Chinese, and other Mongol races, and partly intermixed with them. They possess an ancient literature, and alone among the surrounding peoples have preserved some of the artistic tastes and powers of their ancestors: in painting, sculpture and music they stand high above any people of Mongolian race. They have distinctly Caucasian features, the hair black and often somewhat curled or wavy, while in middle age they are often well bearded though the beard is usually extirpated. They are tall and well made, but are very dark in colour as compared with the Siamese or even the Malays. The modern Khmers do not build or carve in stone like their ancestors, but they imitate the old designs very skilfully in wood. Every article of furniture and all household utensils are more or less ornamented with carved designs; boats, pagodas and houses are carved wherever there is room for such decoration. They also make great figures of gods or of Budha in masonry, wood, or bronze, constructed in several pieces, and having the same general characteristics as those of the old temples.
The two portraits here given (Fig. 79) are copied from La Royaume de Cambodge by J. Moura, and are stated to be from photographs of two Cambodians, one being a native doctor, and although very coarsely engraved they well show the marked Caucasian features. In the Atlas to Garnier's work there are some coloured portraits of these people, as well as of savages from the forests and mountains of the interior, which have even more pronounced European features. It seems therefore not improbable that the civilized Indan type, which invaded
VOL. I. I I
and conquered the country more than four hundred years before the Christian era, found it inhabited by two races, one dark and more allied to themselves, others lighter coloured of Mongolian type. The former would probably become partially civilized and intermingled with their conquerors, while both may have been trained to constructive work under the teaching of the superior race. The wide area over which the ruins are spread, as well as

FIG. 79.—MODERN CAMBODIANS OR KHMERS.
the enormous labour that must have been expended on them, show that the whole area of the south-eastern Asiatic peninsula must have been densely peopled by a civilized nation, although consisting of distinct tribes only partially amalgamated. It is interesting to note that some of the sculptured figures in the ruins bear a considerable resemblance to the existing Khmers (Fig. 80). The radical difference from the Mongolian races around them is also indicated in the Khmer language, which
differs fundamentally from all the languages from Burma to China in being polysyllabic, in the absence of intonation, and in grammatical structure.
We thus see that for a period of more than two thousand years there was, in the great south-eastern peninsula of Asia, a dense population of various races ruled over by a highly civilized superior race of undoubted Caucasian type. But there are still existing the remains of several other tribes, now in a savage state, which are

FIG. 80.—SCULPTURES FROM THE RUINED CITIES.
more or less closely allied to these. Such are the two figured in Garnier's Atlas as inhabiting the mountains of Se-don and the valley of Se-cong, both having fine aquiline noses, good features, and long wavy hair with a dark brown skin; while the Chreans inhabiting the frontiers of Cambodia and Annam are light in colour but with equally Caucasian features. The Khayems near the sources of the Irawaddi have already been mentioned as Caucasic; and I have been recently informed by a gentleman in the service of the Chinese government that, on
I I 2
the western frontier he has seen very fair people with European features and a quite distinctive costume, especially a peculiar head dress closely resembling in its form the Grecian helmet, and quite unlike anything used in other parts of the East. It is therefore highly probable that at some remote period the whole of eastern and south-eastern Asia was inhabited by Caucasian races in various stages of development, before the incursions of the more hardy Mongols of the north broke them up and largely exterminated them.
For more than a quarter of a century it has been recognised by anthropologists that some of the savage and semi-civilised peoples of southern and eastern Asia really belong to the Caucasian type. One of the most interesting of these isolated fragments is the curious hairy race called Ainos, inhabiting the island of Yezo in Japan, and the southern part of Sakhalin, now belonging to Russia. They are believed to be the aborigines of the country, and to have once spread over the whole of the islands, being the barbarians found there by the early Japanese invaders who exterminated them from the more southern and fertile islands. They are a fine people physically and of a mild disposition, but very low as regards material civilization, and extremely conservative of their old customs and mode of life. Mrs. Bishop describes them as about the middle height, broad-chested, the arms and legs short and muscular, the hands and feet large. The bodies of many of them are covered with short bristly hair, as shown in the portrait on p. 467. The forehead is high, broad, and prominent, giving the appearance of intellectual power. The nose is straight but short, the cheek-bones not high. The eyes are large, rather deep set, and very beautiful owing to their soft expression and rich brown colour; and the skin is of an Italian olive tint, and light enough to show the changes of colour in the cheeks. In character they are truthful; infanticide is unknown, and aged parents receive filial reverence, kindness, and support, while in their social and domestic relations there is much to admire. They are treated as barbarians and inferiors by their Japanese rulers, yet it is impossible
to avoid a suspicion that they are fully the equals of their masters, when we see the refinement and beauty of their

FIG. 81.—AINO MAN, URAP.
best representatives as shown in the photograph here reproduced (Fig. 81), a type which is very similar to the man figured, with his wife, in Captain Marshall's
Phrenologist among the Todas—a tribe in southern India —under the title "Adam and Eve."
The fact that a complete state of savagery is quite compatible with elevation of type is shown by the Veddahs of Ceylon, believed to be the aborigines of the island and

FIG. 82.—A VEDDAH SHOOTING GAME.
still living in the dense forests in a completely savage state. They are looked down upon by the other inhabitants as little removed from wild beasts, yet their physical characteristics show them to be a fine race, with all the essential characteristics of the Caucasian type. The reproductions of three very beautiful photographs of these interesting people (Figs. 82, 83, and 84), show them

FIG. 83.—A GROUP OF VEDDAHS.

FIG. 84.—A VEDDAH HUNTER (SAME AS IN FIG. 82).
to be the equals of the Ainos and Todas in perfection of form and features, and to be unmistakably of the same fundamental type as ourselves.
Probable Origin of the Brown Polynesians.
We are now in a position to explain, with some probability, the presence of so high a type (of undoubted Caucasian affinities) in the Pacific, from the Carolines to Easter Island and from Hawaii to New Zealand. At a time when the whole eastern coast of tropical and subtropical Asia was occupied by varieties of this race, which extended also to Japan and to its outlying islands, canoes would be occasionally carried out to sea by tempests, and in this way population would spread farther and farther from the continent. Taking Japan as a probable starting point, owing to the enormous extent of its coast line and its inland seas and straits leading to the development of bold navigators, first the Bonin Islands, then the Ladrones and the Carolines would be thus peopled, since for at least half the year north-westerly winds and gales are prevalent between Japan and the northern tropic. Having once reached thus far, slowly but surely, perhaps after hundreds of years, when these small and unvaried islands became overpopulated, further eastward migration would occur, either by accident or design, to the Marshall group, whence an almost continuous series of groups and islets extends south-eastward through the Gilbert and Ellice Islands to Tonga and Samoa, the first large mountainous and fertile islands in this direction adapted to supply all the conditions for an enjoyable existence. These last formed the centre from which, at a later period when they had become fully peopled, the race spread to the Sandwich Islands and to New Zealand.
This migration from temperate Asia will avoid all the difficulties of the ordinary view of an origin from the Malay Islands, since in them there is no indication of an early occupation by a Caucasian race, while the distances to be traversed are far greater, and there are on the route abundance of islands, thinly inhabited either by people
of Papuan or Malayan race, among whom the primitive Caucasians could have settled; and, if they ultimately passed on eastward, they would certainly have carried with them some indication of such a residence in their language, in the use of the bow-and-arrow, and in the art of making pottery. The deficiency of the whole of the Mahoris in this latter particular is well explained by the theory of their migration step by step through Micronesia from the coasts of Japan or China. In these small islands, mostly of coral with occasionally some basaltic rock, the materials for making pottery would rarely exist, even if among the immigrants there were any persons who knew how to make it. And as it might be a dozen generations, or even much more, before their descendants reached such large islands as those of the Tonga or Samoa groups, all idea of such utensils would have long died out, and the people would be quite content with the modes of cookery they had learnt during their long eastward migration.
One other outlier of these brown Caucasians is found in the Mentawi islands, the southernmost of the long chain that lies off the west coast of Sumatra. These people are described as being like Polynesians, but unlike all the true Malayan tribes. Their language in its abundance of vowels is like that of the Mahoris, and the people resemble them also in the mildness of their disposition and their love of floral decoration. Professor Keane regards them as the last remnant in this direction of the ancient Caucasic people that once occupied southeastern Asia and perhaps what are now the larger Malay Islands, but who were elsewhere driven out or exterminated by the Mongolian invasion.
Comparison of Australians and the Lower Caucasians.
Returning now to the Australians let us see how far they will bear comparison with the remote outposts of the Caucasian race, as illustrated by the reproductions of a few of Professor Spencer's beautiful photographs of the Central Australian tribes. The man of the Arunta tribe (Fig. 85), with his fine beard, his wavy locks and good-
looking pleasant face, though a little coarser is not very far removed from the man of Samoa (at p. 413), or even from the Tonga man (at p. 411), the very handsomest I have

FIG. 85.—ARUNTA TRIBE. MAN WITH FINE BEARD.
yet seen a portrait of. The two old men guarding the sacred "Churinga" (Fig. 86) are hardly less good looking; but I have nowhere been able to obtain a good portrait of a young Australian of pure type and in the prime of life.

FIG. 86.—OLD MEN WITH CHURINGA—SACRED IMPLEMENTS OR "BULL-ROARERS."
The nearest approach is the accompanying portrait of a native of the Northern Territory, from Admiral Maclear's collection (Fig. 87). This shows us a similar type to that of the old man figured at p. 462, and the lank slightly wavy hair indicates that there is no admixture of Papuan.

FIG. 87.—NATIVE OF THE NORTHERN TERRITORY, AUSTRALIA.
The face is very similar to that of some of the Hill tribes of the Indian peninsula—the so-called Dravidians—most of whom are probably Caucasians of low type.
Again, the two Australian women here represented (Figs. 88 and 89) are but little inferior in type to the woman of Samoa (at p. 414), or even to the Hawaiian girl
(at p. 409). In all these cases the Australians have somewhat coarser features of a slightly more negroid type, but

FIG. 88.—WOMAN AND CHILD. ARUNTA TRIBE.
in essential characteristics they agree fairly well. The Queensland woman (p. 463) is so much more negroid that
FIG. 89.—YOUNG WOMAN. ARUNTA TRIBE.
she most likely has some ancestral cross of Papuan, Negrito, or even true negro blood, since during the last century negro slaves were to be found on many European ships.
From the existence of a different and very low type in Tasmania, apparently allied to the Negrito more than to the Papuan, it seems probable that they may have been the primitive inhabitants of Australia or some portion of it, and that they were partly absorbed partly exterminated by the Australians, who themselves must have entered the country at a very remote epoch. Such an intermixture might explain the coarseness of features which removes the Australians so far from all other remnants of the Caucasian type, with which they yet, in their best examples, unmistakably accord. Our general conclusion is, therefore, that both Polynesians and Australians belong to the same fundamental division of the human race as ourselves—the Caucasian. The Polynesians are considerably advanced intellectually and of a higher type physically, and, although somewhat coarser in features, are in both respects the equals of average Europeans. The Australians are far coarser physically and lower mentally and morally. They are distinctly nearer to the animal progenitors of man, and must be of immense antiquity in their present abode. Yet, as they possess the main features of the Caucasian type very well marked, they carry us back but a very few steps towards the primitive man before he was differentiated into the three great races—the Mongoloid, Negroid, and Caucasian.
CHAPTER XXII
THE PROBLEM OF INSTINCT1
THERE is probably no subject in the whole range of biology, the study of which has been so universally neglected as Instinct. Both scientific and popular writers continually refer to it as if its nature and limitations were matter of common knowledge, and its facts so well established as to be almost above criticism. Yet when we ask how it is known that certain actions of man or animals are due to instinct and not to experience or imitation, we find an almost total absence of accurate observation or experiment, while hardly two writers are agreed as to the exact meaning of the term. It is only within the last quarter of a century that a few biologists have made any careful experiments on the phenomena presented by the actions of the higher animals, under such conditions as entirely to exclude the agency of imitation or of parental guidance; and although these experiments are as yet quite insufficient in quantity and far too limited in scope, having regard to the wide field covered by the actions and behaviour usually considered to be instinctive, yet the results reached are already very interesting, and are sufficient to show us that we need not despair of a complete solution of the problem, at all events as regards the higher animals.
One of the first English observers to attack the problem
1 Habit and Instinct, by C. Lloyd Morgan, F.G.S. Pp. 351. London: Edward Arnold, 1896. Price 16s.
VOL. I. K K
on the experimental method was the late Mr. Douglas Spalding, who in 1873, in Macmillan's Magazine, described a number of experiments on young chicks, and ducklings, carefully blinded for the first few hours after birth. His conclusions were, that these young birds not only showed intuitive power of walking, scratching, and pecking, but also possessed intuitive knowledge—or acted as if they possessed such knowledge—of various kinds. He asserted that they were afraid of bees and of the cry of a hawk, and that they intuitively knew the meaning of a hen's call-note and danger-signal when heard for the first time. These results were opposed to Mr. Spalding's preconceived ideas, and were therefore the more readily accepted by naturalists, and they have been frequently quoted as settling this point—the possession by young animals of instinctive knowledge as well as the power of co-ordinated movements of various kinds. Now, Professor Lloyd Morgan has repeated all these experiments many times and with a considerable variety of species, and, while confirming many of Mr. Spalding's observations and conclusions, has shown that those just referred to are erroneous. More important still, he has shown exactly where and why the conclusions arrived at are erroneous, and has thus afforded most valuable guidance to future experimenters in this interesting inquiry. Some examples of these corrections are the following.
Mr. Spalding noticed a difference in the behaviour of young chicks to flies and to bees, and concluded that they "gave evidence of instinctive fear of the sting-bearing insects." This, if true, would be very important, since it would show an intuitive knowledge of the dangerous character of a special insect, and if of one, presumably of all the more common dangerous or hurtful objects, antecedent to experience. But the whole series of observations made by Professor Morgan himself, as well as those made by other good observers, shows that young chicks have no such perception of the qualities of objects. They pick up stones as well as grain, bits of red worsted as well as worms, gaudy-coloured inedible caterpillars
as well as those which are edible. They do not recognize water till they have felt it, and they do not know that water is drinkable till contact with the beaks sets up the nervous and muscular reactions of drinking. By a series of careful experiments Professor Lloyd Morgan shows that young chicks have no fear of bees as bees, but merely because they are large and unusual. They are equally suspicious of a large fly or beetle, and, though eating small worms greedily, are afraid of a large one. And when the chicks were a few days old, and were no longer afraid of large flies, they showed no fear even of wasps, when presented to them for the first time.
Very similar is the correction of Spalding's statements that his young chicks gave the danger-signal when a hawk was flying high overhead, and that a young turkey showed equal alarm when a young hawk, kept in a cupboard, uttered a shrill sound; whence he concluded that the fear of birds of prey, whether seen or heard, was instinctive. For it is now shown that any strange object or any unusual sound causes exactly similar alarm when first seen or heard by any kind of young birds, and Mr. Hudson, of La Plata fame, came to a similar conclusion. Other cases which have been thought to prove instinctive dread of enemies in various young animals are shown to be explicable on similar principles; any sight, or sound or smell, very different from what they have been accustomed to, alarms them, and they learn what are really dangerous either through the actions of their parents or by their own personal experience.
But, though young birds and mammals do not possess instincts which enable them to discriminate between objects that may be useful and those that may be hurtful to them, they often possess the most wonderful acuteness of the senses and powers of co-ordinated muscular action, which enable them rapidly to acquire the knowledge that is essential to them. The young chick only a few hours out of the shell walks and runs, and is able to pick up small objects in its beak, some being rejected and others swallowed. The young duck swims when put into the
K K 2
water, or when it accidentally walks into it, but it has no instinctive desire for it, and does not, as is often stated, run to it from a distance. Young dippers dive perfectly the first time they reach the water, and young swallows fly with great precision, avoiding obstacles almost as readily as do the old birds. With such congenital powers, and with an instinctive fear or suspicion of everything that is strange to them, they learn with marvellous rapidity; and having once found that a particular object is disagreeable or unfit for food, they rarely require a second lesson, and thus in a few days accumulate a stock of experience, which, unless the process has been closely watched, may easily be set down to instinct.
About one-third part of Professor Lloyd Morgan's work is devoted to such experiments and observations on young birds and mammals as have now been indicated, and the amount of new and varied information here brought together is sufficiently large to form the basis of sound reasoning on the nature and limitations of the faculties involved; and perhaps no living biologist is better fitted to do this successfully than the author. In the series of chapters headed: "The Relation of Consciousness to Instinctive Behaviour," "Intelligence and the Acquisition of Habits," "Imitation," and "The Emotions in Relation to Instinct," we have a careful and interesting study of the physiological and psychological aspects of the facts that have been laid before us; a study which is in the highest degree instructive, and which will serve to guide future students of the subject both as to the interpretation of the facts already established, and as to the observations most needed for the elucidation of matters which are still unsettled. These chapters, however, are hardly suited for illustration 01 summary, and we will therefore pass on to those which deal with the alleged instincts of adult animals, and with some of the most disputed questions which now divide biologists; but, before doing so, it will be well to quote the author's definition of instinct, as well as the conclusions he has reached as to its nature.
At the end of the first chapter, which gives a popular sketch of the facts which demand explanation or verification, Professor Morgan says:—
"We may now sum up what has been advanced in the foregoing discussion, and say that, from the biological point of view, instincts are congenital, adaptive, and co-ordinated activities of relative complexity, and involving the behaviour of the organism as a whole. They are not characteristic of individuals as such, but are similarly performed by all like members of the same more or less restricted group, under circumstances which are either of frequent recurrence or are vitally essential to the continuance of the race. … They are to be distinguished from habits which owe their definiteness to individual acquisition and the repetition of individual performance."
And after having described the various actions of young birds antecedent to experience, our author summarizes some of the conclusions to be drawn from the observations as follows:—
"1. That which is congenitally definite as instinctive behaviour is essentially a motor response or train of motor responses. Mr. Herbert Spencer's description of instinct as compound reflex action is thus justified.
2. These often show very accurate and nicely adjusted congenital or hereditary co-ordinations.
3. They are evoked by stimuli, the general type of which is fairly definite, and may, in some cases, be in response to particular objects. Of the latter possibility we have, however, but little satisfactory evidence.
4. There does not seem to be any convincing evidence of inherited ideas or knowledge (as the term is popularly used); that is to say, the facts can be equally well explained on the view that what is inherited is of the nature of an organic response.
5. Association of ideas is strong, and is rapidly formed as the result of individual acquisition.
6. Acquired definiteness is built, through association, on the foundation of congenital responses, which are modified, under experience, to meet new circumstances.
7. Acquired definiteness may pass, through frequent repetition, into more or less stereotyped habit."
Having thus given the author's standpoint and his main conclusions from a body of well-observed facts, we will pass on to his treatment of those activities of adult animals which are generally classed as instinctive.
In the chapter on "Some Habits and Instincts of the Pairing Season," the songs, dances, and displays of plumage by birds are described, and Professor Lloyd Morgan seems inclined to the conclusion that their great development indicates the action of that form of sexual selection termed "preferential mating." Some additional observations are quoted which support this view, but the final conclusion is that—"in all these matters further and fuller evidence from direct observation is to be desired."
The next chapter is on "Nest-building, Incubation, and Migration," and affords much matter for careful consideration. As to nest-building the present writer has always urged that careful experiments are required before we can accept as instinctive the building of a peculiar type of nest by each species of bird; and we find a few such recent experiments now adduced. But when we remember how such a careful experimenter as Spalding was led to wrong conclusions through not varying his experiments sufficiently, the few experiments yet made on nest-building under conditions by no means rigid and with results not described in sufficient detail, can hardly be accepted as settling the question. This is one of the problems that can only be finally settled by experiments tried on a large scale and with every precaution, and the results preserved for comparison and study; and if ever an experimental biological farm is established this subject of nidification would form one of its most valuable and comparatively easy inquiries.
The Migratory Instinct.
Passing over a very interesting discussion as to the habits of the cuckoo and their probable origin, and one hardly less interesting on the habit of the lapwing and many other birds of simulating injury to distract attention from nest or young, we pass on to the broader and more important subject of migration, which, however, is rather briefly treated. The evidence now accumulated seems to justify Professor Lloyd Morgan's conclusion, that while the migratory impulse is innate, yet, "the element of traditional guidance may be effectual, in the migration stream
as a whole, in some way that we have hitherto been unable to observe." The chief obstacle to this view consists in the well-known observations of Herr Gätke at Heligoland, that, during the autumn migration, in the case of the great majority of species, the young birds migrate earliest and alone, the adults following considerably later. But admitting, as every one must, the accuracy of Herr Gätke's observations, does the conclusion necessarily follow? He himself assures us that the birds which rest on or pass within sight of Heligoland only form a fraction of the whole of the migrating hordes, most of them travelling by night at great altitudes, and very few passing within sight of the island, and of these few only, perhaps, one in ten thousand stopping to rest. The fact that young birds of many species are the first to visit Heligoland every year without exception, may possibly be explained by the fact that, while the older birds which lead the way travel high and go on without stopping, a large number of the young fly lower, and being either fatigued by the long unaccustomed flight or attracted by the sight of the land, descend to this elevated and fertile islet for rest and food. The late Mr. Seebohm, whose extensive journeys to Siberia and in various parts of Europe for the purpose of collecting and studying birds, rendered him an authority on this subject, gives the early migration of young birds on the authority of the Heligoland observers, but does not support it by any observation of his own in the northern regions from which so many of the migrants come. In America, although some writers state that young birds migrate first in autumn, Mr. C. Hart Merriam, of the Department of Agriculture, tells us that this notion is "contrary to the experience of most leading American Ornithologists and to the information collected by the Committee on Migration of the American Ornithologists' Union."1 But if we reject the conclusion based upon the Heligoland facts as not necessarily following from them, we shall find that there is not much difficulty in forming a theory which accounts for the main phenomena, and the
1 Report on Bird Migration in the Mississippi Valley, by W. W. Cooke, p. 13. Footnote.
outlines of such a theory have been very well expressed by Mr. Seebohm himself at the end of the chapter on "Migration" in his Geographical Distribution of the Charadriidœ, in the following passage:—
"The assemblage of migratory birds in large flocks, which in many cases wait for a favourable wind before they venture to cross wide stretches of sea, and consequently start altogether as soon as the weather is suitable, and arrive on the other side in enormous numbers or rushes; the keen sight of birds and their extraordinary memory for locality; the great variety of routes chosen, and the pertinacity with which each species keeps to its own route—these and many other facts all point in one direction. The desire to migrate is a hereditary impulse, to which the descendants of migratory birds are subject in spring and autumn, which has acquired a force almost, if not quite, as irresistible as the hereditary impulse to breed in the spring. On the other hand, the routes of migration have to be learned by individual experience. The theory that the knowledge of when and where to migrate is a mysterious gift of nature, the miraculous quality of which is attempted to be concealed under the semi-scientific term of instinct, is no longer tenable."
The views here expressed appear to me to harmonise well with the general conclusions as to the nature and limitations of instinct arrived at by Professor Lloyd Morgan, and they are enforced by some considerations which writers on this subject usually overlook. The numerous recorded facts of birds returning year after year to build in the same spots as in the preceding year, indicate that most of the spring migrants are old birds. Not only is this the general belief of observers, but it is rendered probable by the known longevity of most birds, and the obvious circumstance that those which have escaped the dangers of the double migration on the first occasion will be more likely to escape in each succeeding year, so long as health and strength continue. The fact that the breeding population of birds in any country does not increase year by year, but, though there are considerable fluctuations, remains on the average constant, proves that there must be an enormous destruction of the young birds, which certainly amount in number to several times as many as the old ones, and it seems probable that this destruction takes place during the two annual migrations, and more especially during the first one in autumn, when
the young birds have had no practice in long continued flight and no experience of the dangers of the sea. If the birds of more than one season live on the average only four or five years more, it follows that only a very small percentage of the enormous annual progeny of young birds can survive to take their place. Hence it may well be that all those countless myriads of birds of the year that visit Heligoland are among the failures which, if they leave the island, perish in the waters. We know that enormous numbers must perish during each year, and where so likely as during that first attempt to traverse the North Sea? This is rendered almost certain by the recently issued Report of the British Association Committee on Bird Migration, in which it is stated that at the various periods of the great autumnal rushes at Heligoland, when countless thousands of birds visit or pass over that island, no corresponding influx has been noticed on our east coasts during the four successive years that the two records have been compared. Either then, these myriads of birds passed southward to Holland or flew out to sea and were mostly lost. As Herr Gätke does not mention any corresponding flights along the coasts of Holland and Belgium it is to be presumed that they have not been noticed, and we are almost forced to the conclusion that the greater part of these young birds, whose immense numbers at Heligoland excite so much astonishment, are really failures, and form a portion of those which are annually eliminated by the severe test of migration; and if this be so, much of the marvel supposed to attach to the successful migration of young birds disappears, since such "successful migration," except in the case of a small percentage, does not occur.
The alleged Homing Instinct.
There is one alleged instinct of great popular interest which Mr. Lloyd Morgan does not deal with—the means by which many of our domestic animals, especially cats, dogs, and horses, find their way home under circumstances which seem to preclude any direct guidance by the senses.
Narratives of the most marvellous character have been published, but, unfortunately, no systematic experiments appear to have been made, except a few by the late Mr. Romanes. These showed that a dog could exactly track its master's footsteps by scent, and it was proved that the scent was derived from his shoes, which must of course be full of perspiration and other emanations from the skin, because when he wore new shoes or those of another person his track could not be followed. In Nature, vol. vii., there is a considerable discussion of this subject, and many remarkable cases are narrated by various correspondents, but in none are all the data given for arriving at a rational conclusion on the question. I then wrote summarizing the discussion (Nature, vol. viii., p. 65), and suggested a series of experiments, which would give us the real information required. My suggestion was that a dog whose antecedents were known should be taken by a circuitous route by rail and road to some spot where he had never been before, and should be there handed to some person he did not know, who should carry him a short distance, and on releasing him should keep him in sight, either on foot or horseback, till the animal returned home, noting carefully every movement and action. A moderate number of such experiments would settle the question of instinct or sense-observation, and it is to be hoped, now that a more intelligent interest is taken in the subject, such experiments will be made.
Theoretically, any instinct of direction is almost inconceivable, because quadrupeds in a state of nature do not require such an instinct. They learn, step by step, the surroundings of their birth-place, extending their range, perhaps, year by year, but never requiring to go back over a country they have not previously traversed. In their case ordinary memory, assisted by very acute senses, would be all they would need. In the case of domestic animals returning home, we find that the recorded time elapsed varies from a few hours to several days or weeks, even when the distance can be easily traversed in a day. This shows that an instinctive sense of direction cannot always be the agency employed; and, as in no one case is the
exact route of the animal's return known, the assertion so often made, that a special sense of direction is required to explain the facts, cannot be justified.
Are Acquired Characters Inherited?
Two very interesting chapters, near the end of the volume, are those entitled "Are Acquired Characters Inherited," and "Modification and Variation." In the first of these the bearing of the whole of the phenomena of instinct on the vexed question of inheritance is pointed out, and the conclusion is reached, that although there is little or no satisfactory evidence of the transmission of acquired modifications—that is of habits, or their effects on the organism, as opposed to instincts, yet there are many curious facts which seem to indicate some connection between congenital and acquired characters. What this connection is, the chapter on Modification and Variation attempts to show.
Modification of the individual by the environment, whether in the direction of structure or of habits, is universal and of considerable amount, and it is almost always, under the actual conditions, a beneficial modification. But every kind of beneficial modification is also being constantly effected through variation and natural selection, so that the beautifully perfect adaptations we see in nature are the result of a double process, being partly congenital, partly acquired. Acquired modification thus helps on congenital change by giving time for the necessary variations in many directions to be selected, and we have here another answer to the supposed difficulty as to the necessity of many coincident variations in order to bring about any effective advance of the organism. In one year favourable variations of one kind are selected and individual modifications in other directions enable them to be utilised: in Professor Lloyd Morgan's words—"Modification as such is not inherited, but is the condition under which congenital variations are favoured and given time to get a hold on the organism, and are thus enabled by degrees to reach the fully adaptive level." The same result
will be produced by Professor Weismann's recent suggestion of "germinal selection," so that it now appears as if all the theoretical objections to the "adequacy of natural selection" have been theoretically answered.
Biologists owe a debt of gratitude to Professor Lloyd Morgan for this most interesting and suggestive volume. It exhibits all the clearness and philosophical acumen which characterize the writings of the author, and although in his desire to be impartial he has sometimes suggested difficulties which are more apparent than real, yet the work is on the whole an admirable introduction to the study of a most important and fascinating branch of biology, now for the first time based upon a substantial foundation of carefully observed facts and logical induction from them.
CHAPTER XXIII
HUMAN SELECTION
IN one of my latest conversations with Darwin he expressed himself very gloomily on the future of humanity, on the ground that in our modern civilization natural selection had no play, and the fittest did not survive. Those who succeed in the race for wealth are by no means the best or the most intelligent, and it is notorious that our population is more largely renewed in each generation from the lower than from the middle and upper classes. As a recent American writer well puts it, "We behold the melancholy spectacle of the renewal of the great mass of society from the lowest classes, the highest classes to a great extent either not marrying or not having children. The floating population is always the scum, and yet the stream of life is largely renewed from this source. Such a state of affairs, sufficiently dangerous in any society, is simply suicidal in the democratic civilization of our day."1
That the check to progress here indicated is a real one few will deny, and the problem is evidently felt to be one of vital importance, since it has attracted the attention of some of our most thoughtful writers, and has quite recently furnished the theme for a perfect flood of articles in our best periodicals. I propose here to consider very briefly the various suggestions made by these writers; and afterwards shall endeavour to show that when the course of social evolution shall have led to a more rational organization of society, the problem will receive its final solution
1 Hiram M. Stanley in the Arena for June, 1890.
by the action of physiological and social agencies, and in perfect harmony with the highest interests of humanity.
Are the Results of Training Hereditary?
Before discussing the question itself it will be well to consider whether there are in fact any other agencies than some form of selection to be relied on. It has been generally accepted hitherto that such beneficial influences as education, hygiene, and social refinement had a cumulative action, and would of themselves lead to a steady improvement of all civilized races. This view rested on the belief that whatever improvement was effected in individuals was transmitted to their progeny, and that it would be thus possible to effect a continuous advance in physical, moral, and intellectual qualities without any selection of the better or elimination of the inferior types. But of late years grave doubts have been thrown on this view, owing chiefly to the researches of Galton and Weismann as to the fundamental causes to which heredity is due. The balance of opinion among physiologists now seems to be against the heredity of any qualities acquired by the individual after birth, in which case the question we are discussing will be much simplified, since we shall be limited to some form of selection as the only possible means of improving the race.
In order to make the difference between the two theories clear to those who may not have followed the recent discussions on the subject, an illustration may be useful. Let us suppose two persons, each striving to produce two distinct types of horse—the cart-horse and the racer—from the wild prairie horses of America, and that one of them believes in the influence of food and training, the other in selection. Each has a lot of a hundred horses to begin with, as nearly as possible alike in quality. The one who trusts to selection at once divides his horses into two lots, the one stronger and heavier, the other lighter and more active, and, breeding from these, continually selects, for the parents of the succeeding generation, those which most nearly approach
the two types required. In this way it is perfectly certain that in a comparatively short period—thirty or forty years perhaps—he would be able to produce two very distinct forms, the one a very fair racehorse, the other an equally good specimen of a cart-horse; and he could do this without subjecting the two strains to any difference of food or training, since it is by selection alone that our various breeds of domestic animals have in most cases been produced.
On the other hand, the person who undertook to produce similar results by food and training alone, without allowing selection to have any part in the process, would have to act in a very different manner. He should first divide his horses into two lots as nearly as possible identical in all points, and thereafter subject the one lot to daily exercise in drawing loads at a slow pace, the other lot to equally constant exercise in running, and he might also supply them with different kinds of food if he thought it calculated to aid in producing the required effect. In each successive generation he must make no selection of the swiftest or the strongest, but must either keep the whole progeny of each lot, or carefully choose an average sample of each to be again subjected to the same discipline. It is quite certain that the very different kinds of exercise would have some effect on the individuals so trained, enlarging and strengthening a different set of muscles in each, and if this effect were transmitted to the offspring then there ought to be in this case also a steady advance towards the racer and the cart-horse type. Such an experiment, however, has never been tried, and we cannot therefore say positively what would be the result; but those who accept the theory of the non-heredity of acquired characters would predict with confidence that after thirty or forty generations of training without selection, the last two lots of colts would have made little or no advance towards the two types required, but would be practically indistinguishable.
It is exceedingly difficult to find any actual cases to illustrate this point, since either natural or artificial selection has almost always been present. The apparent effects of disuse in causing the diminution of certain
organs, such as the reduced wings of some birds in oceanic islands and the very small or aborted eyes of some of the animals inhabiting extensive caverns, can be as well explained by the withdrawal of the cumulative agency of natural selection and by economy of growth, as by the direct effects of disuse. The following facts, however, seem to show that special skill derived from practice, when continued for several generations, is not inherited, and does not therefore tend to increase. The wonderful skill of most of the North American Indians in following a trail by indications quite imperceptible to the ordinary European has been dwelt upon by many writers, but it is now admitted that the white trappers equal and often excel them, though these trappers have in almost every case acquired their skill in a comparatively short period, without any of the inherited experience supposed to belong to the Indian. Again, for many generations a considerable proportion of the male population of Switzerland has practised rifle-shooting as a national sport, yet in international contests they show no marked superiority over our riflemen, who are, in a large proportion, the sons of men who never handled a gun. Another case is afforded by the upper classes of this country who for many generations have been educated at the universities, and have had their classical and mathematical abilities developed to the fullest extent by rivalry for honours. Yet now, that for some years these institutions have been opened to dissenters whose parents usually for many generations have had no such training, it is found that these dissenters carry off their full share or even more than their share of honours. We thus see that the theory of the non-heredity of acquired characters, whether physical or mental, is supported by a considerable number of facts, while few if any are directly opposed to it. We therefore propose to neglect the influence of education and habit as possible factors in the improvement of our race, and to confine our argument entirely to the possibility of improvement by some form of selection.1
1 Those who desire more information on this subject should read Weismann's Essays on Heredity.
Proposals for the Improvement of the Race.
Among the modern writers who have dealt with this question the opinions of Mr. Galton are entitled to be first considered, because he has studied the whole subject of human faculty in the most thorough manner, and has perhaps thrown more light upon it than any other writer. The method of selection by which he has suggested that our race may be improved is to be brought into action by means of a system of marks for family merit, both as to health, intellect, and morals, those individuals who stand high in these respects being encouraged to marry early by state endowments sufficient to enable the young couples to make a start in life. Of all the proposals that have been made tending to the systematic improvement of our race, this is one of the least objectionable, but it is also I fear among the least effective. Its tendency would undoubtedly be to increase the number and to raise the standard of our highest and best men, but it would at the same time leave the bulk of the population unaffected, and would but slightly diminish the rate at which the lower types tend to supplant or to take the place of the higher. What we want is, not a higher standard of perfection in the few, but a higher average, and this can best be produced by the elimination of the lowest of all and a free intermingling of the rest.
Something of this kind is proposed by Mr. Hiram M. Stanley in his article on "Our Civilization and the Marriage Problem," already referred to. This writer believes that civilizations perish because, as wealth and art increase, corruption creeps in, and the new generations fail in the work of progress because the renewal of individuals is left chiefly to the unfit. The two great factors which secure perfection in each animal race—sexual selection by which the fit are born, and natural selection by which the fittest survive—both fail in the case of mankind, among whom are hosts of individuals which in any other class of beings would never have been born, or, if born would never survive. He argues that,
VOL. I. L L
unless some effective measures are soon adopted and strictly enforced, our case will be irremediable; and, since natural selection fails so largely, recourse must be had to artificial selection. "The drunkard, the criminal, the diseased, the morally weak should never come into society. Not reform but prevention should be the cry." The method by which this is proposed to be done is hinted at in the following passages: "In the true golden age, which lies not behind but before us, the privilege of parentage will be esteemed an honour for the comparatively few, and no child will be born who is not only sound in body and mind, but also above the average as to natural ability and moral force"—and again—"The most important matter in society, the inherent quality of the members which compose it, should be regulated by trained specialists."
Of this proposal and all of the same character we may say, that nothing can possibly be more objectionable, even if we admit that they might be effectual in securing the object aimed at. But even this is more than doubtful; and it is quite certain that any such interference with personal freedom in matters so deeply affecting individual happiness will never be adopted by the majority of any nation, or if adopted would never be submitted to by the minority without a life-and-death struggle.
Another popular writer of the greatest ability and originality, who has recently given us his solution of the problem, is Mr. Grant. Allen. His suggestion is in some respects the very reverse of the last, yet it is, if possible, even more objectionable. Instead of any interference with personal freedom he proposes the entire abolition of legal restrictions as to marriage, which is to be a free contract, to last only so long as either party desires. This alone, however, would have no effect on race-improvement, except probably a prejudicial one. The essential part of his method is, that girls should be taught both by direct education and by the influence of public opinion, that the duty of all healthy and intellectual women is to be the mothers of as many and as perfect children as possible. For this purpose they are re-
commended to choose as temporary husbands the finest, healthiest, and most intellectual men, thus ensuring a variety of combinations of parental qualities which would lead to the production of offspring of the highest possible character and to the continual advancement of the race.1
I think I have fairly summarized the essence of Mr. Grant Allen's proposal, which, though enforced with all his literary skill and piquancy of illustration, can, in my opinion, only be fitly described by the term already applied to it by one of his reviewers, "detestable." It purports to be advanced in the interests of the children and of the race; but it would necessarily impair that family life and parental affection which are the prime essentials to the well-being of children; while, though it need not necessarily produce, it would certainly favour, the increase of pure sensualism, the most degrading and most fatal of all the qualities that tend to the deterioration of races and the downfall of nations. One of the modern American advocates of greater liberty of divorce, in the interest of marriage itself, thus admirably summarizes the essential characteristics and purport of true marriage. "In a true relation, the chief object is the loving companionship of man and woman, their capacity for mutual help and happiness, and for the development of all that is noblest in each other. The second object is the building up a home and family, a place of rest, peace, security, in which child-life can bud and blossom like flowers in the sunshine."2 For such rest, peace, and security, permanence is essential. This permanence need not be attained by rigid law, but by the influence of public opinion, and, more surely still, by those deep-seated feelings and emotions which, under favourable conditions, render the marriage tie stronger and its influence more beneficial the longer it endures. To me it appears that no system of the relations of men and women could be more fatal to the happiness of individuals, the well-being
1 See "The Girl of the Future" in The Universal Review, May, 1890, and a previous article entitled, "Plain Words on the Woman Question," in The Fortnightly Review, October, 1889.
2 Elizabeth Cady Stanton, in the Arena, April, 1890.
L L 2
of children, or the advancement of the race, than that proposed by Mr. Grant Allen.
Objections to all the preceding Proposals.
Before proceeding further with the main question it is necessary to point out that, besides the special objections to each of the proposals here noticed, there is a general and fundamental objection. They all attempt to deal at once, and by direct legislative enactment, with the most important and most vital of all human relations, regardless of the fact that our present phase of social development is not only extremely imperfect, but vicious and rotten at the core. How can it be possible to determine and settle the relations of women to men which shall be best alike for individuals and for the race, in a society in which a very large proportion of women are obliged to work long hours daily for the barest subsistence, while another large proportion are forced into more or less uncongenial marriages as the only means of securing some amount of personal independence or physical well-being? Let any one consider, on the one hand, the lives of the wealthy as portrayed in the society newspapers, and even in the advertisements of such papers as The Field and The Queen, with their endless round of pleasure and luxury, their almost inconceivable wastefulness and extravagance, indicated by the cost of female dress and such facts as the expenditure of a thousand pounds on the flowers for a single entertainment; and, on the other hand, the terrible condition of millions of workers—men, women, and children—as detailed in the Report of the Lords' Commission on Sweating, on absolutely incontestable evidence, and the still more awful condition of those who seek work of any kind in vain, and, seeing their children slowly dying of starvation, are driven in utter helplessness and despair to murder and suicide. Can any thoughtful person admit for a moment that, in a society so constituted that these overwhelming contrasts of luxury and privation are looked upon as necessities, and are treated by the Legislature as matters with which it has
practically nothing to do, there is the smallest probability that we can deal successfully with such tremendous social problems as those which involve the marriage tie and the family relation as a means of promoting the physical and moral advancement of the race? What a mockery to still further whiten the sepulchre of modern society, in which is hidden "all manner of corruption," with schemes for the moral and physical advancement of the race!
Social Advance will result in Improvement of Character.
It is my firm conviction, for reasons which I shall state presently, that, when we have cleansed the Augean stable of our existing social organization, and have made such arrangements that all shall contribute their share of either physical or mental labour, and that all workers shall reap the full and equal reward of their work, the future of the race will be ensured by those laws of human development that have led to the slow but continuous advance in the higher qualities of human nature. When men and women are alike free to follow their best impulses; when idleness and vicious or useless luxury on the one hand, oppressive labour and starvation on the other, are alike unknown; when all receive the best and most thorough education that the state of civilization and knowledge at the time will admit; when the standard of public opinion is set by the wisest and the best, and that standard is systematically inculcated on the young; then we shall find that a system of selection will come spontaneously into action which will steadily tend to eliminate the lower and more degraded types of man, and thus continuously raise the average standard of the race. I therefore strongly protest against any attempt to deal with this great question by legal enactments in our present state of unfitness and ignorance, or by endeavouring to modify public opinion as to the beneficial character of monogamy and permanence in marriage. That the existing popular opinion is the true one is well and briefly shown by Miss Chapman in a recent number of Lippincott's Magazine; and as her statement of the case expresses my
own views, and will, I think, be approved by most thinkers on the subject, I here give it.
"1. Nature plainly indicates permanent marriage as the true human relation. The young of the human pair need parental care and supervision for a great number of years.
2. Instinct is strongly on the side of indissoluble marriage. In proportion as men leave brutedom behind and enter into the fulness of their human heritage, they will cease to tolerate the idea of two or more living partners.
3. History shows conclusively that where divorce has been easy, licentiousness, disorder, and often complete anarchy have prevailed. The history of civilization is the history of advance in monogamy, of the fidelity of one man to one woman, and one woman to one man.
4. Science tells the same tale. Physiology and Hygiene point to temperance, not riot. Sociology shows how man, in spite of himself, is ever striving, through lower forms, upward, to the monogamic relation.
5. Experience demonstrates to every one of us, individually, the superiority of the indissoluble marriage. We know that, speaking broadly, marriages turn out well or ill in proportion as husband and wife are—let me not say loving—but loyal, sinking differences and even grievances for the sake of children and for the sake of example."
We have now to consider what would be the probable effect of a condition of social advancement, the essential characteristics of which have been already hinted at, on the two great problems—the increase of population, and the continuous improvement of the race by some form of selection which we have reason to believe is the only method available. In order to make this clear, however, and in order that we may fully realize the forces that would come into play in a just and rational state of society, such as may certainly be realized in the not distant future, it will be necessary to have a clear conception of its main characteristics. For this purpose, and without committing myself in any way to an approval of all the details of his scheme, I shall make use of Mr. Bellamy's clear and forcible picture of the society of the future, as he supposes it may exist in America in little more than a century hence.1
The essential principle on which society is supposed to be
1 Looking Backward. See specially chapters vii., ix., xii., and xxv.
founded is that of a great family. As in a well-regulated modern family the elders, those who have experience of the labours, the duties, and the responsibilities of life, determine the general mode of living and working, with the fullest consideration for the convenience and real well-being of the younger members, and with a recognition of their essential independence. As in a family, the same comforts and enjoyments are secured to all, and the very idea of making any difference in this respect to those who from mental or physical disability are unable to do so much as others, never occurs to any one, since it is opposed to the essential principles on which a true society of human brotherhood is held to rest. As regards education all have the same advantages, and all receive the fullest and best training, both intellectual and physical; every one is encouraged to follow out those studies or pursuits for which they are best fitted, or for which they exhibit the strongest inclination. This education, the complete and thorough training for a life of usefulness and enjoyment, continues in both sexes till the age of twenty-one (or thereabouts), when all alike, men and women, take their place in the lower ranks of the industrial army in which they serve for three years. During the latter years of their education, and during the succeeding three years of industrial service, every opportunity is given them to see and understand every kind of work that is carried on by the community, so that at the end of the term of probation they can choose what department of the public service they prefer to enter. As every one—men, women, and children alike—receive the same amount of public credit—their equal share of the products of the labour of the community, the attractiveness of various pursuits is equalized by differences in the hours of labour, in holidays, or in special privileges attached to the more disagreeable kinds of necessary work, and these are so modified from time to time that the volunteers for every occupation are always about equal to its requirements. The only other essential feature that it is necessary to notice for our present purpose is the system of grades, by which good conduct, perseverance, and intelligence in every department of industry and
occupation are fully recognized, and lead to appointments as foremen, superintendents, or general managers, and ultimately to the highest offices of the state. Every one of these grades and appointments is made public; and as they constitute the only honours and the only differences of rank, with corresponding insignia and privileges, in an otherwise equal body of citizens, they are highly esteemed, and serve as ample inducements to industry and zeal in the public service.
At first sight it may appear that in any state of society whose essential features were at all like those here briefly outlined, all the usual restraints to early marriage as they now exist would be removed, and that a rate of increase of the population unexampled in any previous era would be the result, leading in a few generations to a difficulty in obtaining subsistence, which Malthus has shown to be the inevitable result of the normal rate of increase of mankind when all the positive as well as the preventive checks are removed. As the positive checks—which may be briefly summarized as war, pestilence, and famine—are supposed to be non-existent, what, it may be asked, are the preventive checks which are suggested as being capable of reducing the rate of increase within manageable limits? This very reasonable question I will now endeavour to answer.
Natural Checks to rapid Increase.
The first and most important of the checks upon a too rapid increase of population will be the comparatively late average period of marriage, which will be the natural result of the very conditions of society, and will besides be inculcated during the period of education, and still further enforced by public opinion. As the period of systematic education is supposed to extend to the age of twenty-one, up to which time both the mental and physical powers will be trained and exercised to their fullest capacity, the idea of marriage during this period will rarely be entertained. During the last year of education, however, the subject of marriage will be dwelt upon, in its bearing on individual happiness and on social well-being, in relation to the welfare of the next generation and to the continuous
development of the race. The most careful and deliberate choice of partners for life will be inculcated as the highest social duty; while the young women will be so trained as to look with scorn and loathing on all men who in any way wilfully fail in their duty to society—on idlers and malingerers, on drunkards and liars, on the selfish, the cruel, or the vicious. They will be taught that the happiness of their whole lives will depend on the care and deliberation with which they choose their husbands, and they will be urged to accept no suitor till he has proved himself to be worthy of respect by the place he holds and the character he bears among his fellow-labourers in the public service.
Under social conditions which render every woman absolutely independent, so far as the necessaries and comforts of existence are concerned, surrounded by the charms of family life and the pleasures of society, which will be far greater than anything we now realize when all will possess the refinements derived from the best possible education, and all will be relieved from sordid cares and the struggle for mere existence, is it not in the highest degree probable that marriage will rarely take place till the woman has had three or four years' experience of the world after leaving college—that is, till the age of 25, while it will very frequently be delayed till 30 or upwards? Now Mr. Galton has shown, from the best statistics available, that if we compare women married at 20 with those married at 29, the proportionate fertility is about as 8 to 5. But this difference, large as it is, only represents a portion of the effect on the rate of increase of population caused by a delay in the average period of marriage. For when the age of marriage is delayed the time between successive generations is correspondingly lengthened; while a still further effect is produced by the fact that the greater the average age of marriage the fewer generations are alive at the same time, and it is the combined effect of these three factors that determines the actual rate of increase of the population.1
But there is yet another factor tending to check the
1 See Inquiries into Human Faculty and Its Development, p. 321; and Hereditary Genius, p. 353.
increase of population that would come into play in a society such as we have been considering. In a remarkable essay on the Theory of Population Herbert Spencer has shown, by an elaborate discussion of the phenomena presented by the whole animal kingdom, that the maintenance of the individual and the propagation of the race vary inversely, those species and groups which have the shortest and most uncertain lives producing the greatest number of offspring; in other words, individuation and reproduction are antagonistic. But individuation depends almost entirely on the development and specialization of the nervous system, through which, not only are the several activities and co-ordinations of the various organs carried on, but all advance in instinct, emotion, and intellect is rendered possible. The actual rate of increase in man has been determined by the necessities of the savage state, in which, as in most animal species, it has usually been only just sufficient to maintain a limited average population. But with civilization the average duration of life increases, and the possible increase of population under favourable conditions becomes very great, because fertility is greater than is needed under the new conditions. The advance in civilization as regards the preservation of life has in recent times become so rapid, and the increased development of the nervous system has been limited to so small a portion of the whole population, that no general diminution in fertility has yet occurred. That the facts do, however, accord with the theory is indicated by the common observation that highly intellectual parents do not as a rule have large families, while the most rapid increase occurs in those classes which are engaged in the simpler kinds of manual labour. But in a state of society in which all will have their higher faculties fully cultivated and fully exercised throughout life, a slight general diminution of fertility would at once arise, and this diminution added to that caused by the later average period of marriage would at once bring the rate of increase of population within manageable limits. The same general principle enables us to look forward to that distant future when the world will be fully peopled, in perfect confidence that an
equilibrium between the birth and death rates will then be brought about by a combination of physical and social agencies, and the bugbear of over-population become finally extinct.1
How Natural Selection will improve the Race.
There now only remains for consideration the means by which, in such a society, a continuous improvement of the race could be brought about, on the assumption that for this purpose education is powerless as a direct agency, since its effects are not hereditary, and that some form of selection is an absolute necessity. This improvement I believe will certainly be effected through the agency of female choice in marriage. Let us, therefore, consider how this would probably act.
It will be generally admitted that, although many women now remain unmarried from necessity rather than from choice, there are always a considerable number who feel no strong inclination to marriage, and who accept husbands to secure a subsistence or a home of their own rather than from personal affection or sexual emotion. In a society in which women were all pecuniarily independent, were all fully occupied with public duties and intellectual or social enjoyments, and had nothing to gain by marriage as regards material well-being, we may be sure that the number of the unmarried from choice would largely increase. It would probably come to be considered a degradation for any woman to marry a man she could not both love and esteem, and this feeling would supply ample reasons for either abstaining from marriage altogether or delaying it till a worthy and sympathetic husband was encountered. In man, on the other hand, the passion of love is more general, and usually stronger; and as in such a society as is here postulated there would be no way of gratifying this passion but by marriage, almost every woman would receive offers, and thus a powerful selective agency would rest with the female sex. Under the
1 A Theory of Population deduced from the General Law of Animal Fertility. Republished from the Westminster Review for April, 1852.
system of education and of public opinion here suggested there can be no doubt how this selection would be exercised. The idle and the selfish would be almost universally rejected. The diseased or the weak in intellect would also usually remain unmarried; while those who exhibited any tendency to insanity or to hereditary disease, or who possessed any congenital deformity would in hardly any case find partners, because it would be considered an offence against society to be the means of perpetuating such diseases or imperfections.
We must also take into account a special factor hitherto, I believe, unnoticed in this connection, that would in all probability intensify the selection thus exercised. It is well known that females are largely in excess of males in our existing population, and this fact, if it were a necessary and permanent one, would tend to weaken the selective agency of women, as it undoubtedly does now. But there is good reason to believe that it will not be a permanent feature of our population. The births indicate a natural tendency in the opposite direction, since they always give a larger proportion of males than females, varying from 3½ to 4 per cent. But boys now die so much more rapidly than girls that when we include all under the age of five the numbers are nearly equal. For the next five years the mortality is nearly the same in both sexes; then that of females preponderates up to 30 years of age, then up to 60 that of men is the larger, while for the rest of life female mortality is again greatest. The general result is that at the ages of most frequent marriage—from 20 to 35—females are between 8 and 9 per cent. in excess of males. But during the ages from 5 to 35 we find a wonderful excess of male deaths from two preventible causes—"accident" and "violence." For the year 1888 the deaths from these causes in England and Wales was as follows:—
Males (5 to 35 years) 4,158
Females (5 to 35 years) 1,1001
Here we have an excess of male over female deaths in
Annual Report of the Registrar-General, 1888, pp. 106–7.
one year of 3,058, all between the ages of 5 and 35, a very large portion of which is no doubt due to the greater risks run by men and boys in various industrial occupations, in sport, and in war. In a state of society in which the bulk of the population were engaged in industrial work, and were all social equals, it is quite certain that almost all these deaths would be prevented, thus bringing the male population more nearly to an equality with the female. But there are also many unhealthy employments in which men are exclusively or more largely engaged, such as the grinders of Sheffield, and many others; and many more men have their lives shortened by labour in unventilated workshops, to say nothing of the loss of life at sea and in war. When the lives of all its citizens are accounted of equal value to the community, no one will be allowed to suffer from such preventible causes as these; and this will still further reduce the mortality of men as compared with that of women. On the whole, then, it seems highly probable that in the society of the future the superior numbers of males at birth will be maintained throughout life, or, at all events, during what may be termed the marriageable period. This will greatly increase the influence of women in the improvement of the race. Being a minority they will be more sought after, and will have a real choice in marriage, which is rarely the case now. This actual minority being further increased by those who, from the various causes already referred to, abstain from marriage, will cause considerable numbers of men to remain permanently unmarried, and as these will consist very largely, if not almost wholly, of those who are the least perfectly developed either mentally or physically, the constant advance of the race in every good quality will be ensured.
This method of improvement, by elimination of the worst, has many advantages over that of securing the early marriages of the best. In the first place it is the direct instead of the indirect way, for it is more important and more beneficial to society to improve the average of its members by getting rid of the lowest types than by raising the highest a little higher. Exceptionally great
and good men are always produced in sufficient numbers, and have always been so produced in every phase of civilization. We do not need more of these so much as we need less of the weak and the bad. This weeding-out system has been the method of natural selection, by which the animal and vegetable worlds have been improved and developed. The survival of the fittest is really the extinction of the unfit. In nature this occurs perpetually on an enormous scale, because, owing to the rapid increase of most organisms, the unfit which are yearly destroyed form a large proportion of those that are born. Under our hitherto imperfect civilization this wholesome process has been checked as regards mankind; but the check has been the result of the development of the higher attributes of our nature. Humanity—the essentially human emotion—has caused us to save the lives of the weak and suffering, of the maimed or imperfect in mind or body. This has to some extent been antagonistic to physical and even intellectual race-improvement; but it has improved us morally by the continuous development of the characteristic and crowning grace of our human, as distinguished from our animal nature.
In the society of the future this defect will be remedied, not by any diminution of our humanity, but by encouraging the activity of a still higher human characteristic—admiration of all that is beautiful and kindly and self-sacrificing, repugnance to all that is selfish, base, or cruel. When we allow ourselves to be guided by reason, justice, and public spirit in our dealings with our fellow-men, and determine to abolish poverty by recognizing the equal rights of all the citizens of our common land to an equal share of the wealth which all combine to produce—when we have thus solved the lesser problem of a rational social organization adapted to secure the equal well-being of all, then we may safely leave the far greater and deeper problem of the improvement of the race to the cultivated minds and pure instincts of the men, and especially of the Women of the Future.
INDEX
A
Aar, gorge of, how formed, 136
the valley of, 129
Aarschlucht, the, 131
Acquired characters, definition of, 317
results of inheritance of, 315, 318, 507
Adaptations not directly caused by environment, 312
not producible by use-inheritance, 343
Aeppli answered by Prof. James Geikie, 143
Affinities of the Papuans, 459
Agassiz, on ancient glaciers, 61
Ainos and Australians, 465
Airy, Sir G. B., on weakness of earth's crust, 47
Allen, Mr. J. A., on variation, 293
Allied species in same area, 282
American erratics, 74
flowers, rambles after, 206
monkeys, 165
Animals, mode of development of, 289
Anthropoid apes, how to keep alive, 160
Arabis anachoretica and A. alpina, 339
Artemia salina changes in salter water, 339
Ateles, 167
Australians are low Caucasians, 468, 471
affinities of, 461, 496
mental and moral character of, 472
Australian women, 494
Aye-aye, 177
Azores, beetles of, 263
few peculiar, 263
violent storms of, 264
B.
Baboons, 156
Baioñ, ruins of, 478
Bateson on discontinuity, 352
Beccari, Dr., on the Papuans, 455
Beeby, Mr., on intrinsic and extrinsic varieties, 314, 340
Beete-Jukes, on valley formation, 15
Beetles, genera absent from Madeira, 256
disguises of, 194
found at sea, 251
not peculiar to Madeira, 259
of Madeira often wingless, 254
Bellamy's forecast of society, 518
Belt on shell-bearing glacial gravels, 93
Big trees of California, 227, 234
Birds, four toes of, 360
of New Guinea, 423
Bishop, Mrs., on the Ainos, 484
Blandford, W. T., on permanence of oceans, 29
Brachiurus, 171
Bradley, Prof. C. B., on age of Sequoias, 230
Branner, Prof., on narrow gorges, 135
none in Brazil, 136
British erratics, teachings of, 88
plants in N. America, 203
not in N. America, 203
Buckland, Dr., on ancient glaciers, 59
Buffalo grass, 214
Buffon on mutability of species, 345
Buff-tip moth, disguise of, 188
Buprestidæ, disguises of, 195
Butterflies and moths, colours of, 186
Butterfly 400 miles at sea, 252
C.
Calaveras grove, 228
California, flora of, 223
forests of, 224
Callithrix, 173
Campbell, Lord G., on Polynesians, 410
Canaries, spiny plants rare, 309
Caucasian affinities of Australians, 465, 496
Caucasians of S.E. Asia, 484
Caverns in N.W. Australia, 20
Cebus, 170
Centetidæ, distribution of, 279
Cercopithecus, 155
Chamberlain, Mr. T. C., on meteoric theory of earth-formation, 57
on rock-scorings, 68
Chapman, Miss, on true marriage, 518
Character modified by social advance 517
Charpentier on Swiss erratics, 73
Checks to increase of population, 520
Chicks, habits of young, 498
Chimpanzee, 154
Chrysothrix, 173
Coleoptera of Madeira, 250
Colobus, 155
Colorado, canon of, 19
Cook, Captain, on natives of Tonga Islands, 410
Cornus Nuttalii, 226
Cosmopolitan plants in Europe, 202
Comrie, Dr., on the Papuans, 453
Cox valley, 10
Cricketers as illustrating adaptation, 332
Critic of natural selection answered, 305
Crosby, Mr., on oceanic islands, 34
Crust of earth affects gravity, 47
Cunningham, Mr., on origin of horns, 321
Curr on the Australian race, 464, 473
Cyanopica Cooki, distribution of, 268
Cynocephalus, 156
D.
Dana, Prof., on oceanic basins, 28
Darwin, Prof. G., 43
on beetles found at sea, 251
on best criterion of ice-action, 67
on character of the Polynesians, 405
on inaccessible valleys, 11, 14
on origin of horns, 322
on permanence of oceans, 28
on useless specific characters, 387
his theory, what it is, 286
his work, effect of, 347
Darwinism compared with Lamarckism, 350
does not explain origin of life, 288
minor objections to, 364
objections to invalid, 287
opponents of, 286
what it really is, 348
Delaporte on ruins in Cambodia, 477
Delbœuf's law, 374
Deltas prove subsidence, 45
Dermal appendages, origin of, 322
Desor on valleys of disruption, 14
Discontinuity, the theory of, 352
Dogs, peculiar habit of, 320
Dome forms of granite, 22
Doone valley, 2
Douglas fir, 227
E
Earth, internal heat of, 41
Earth's crust, Appendix on, 57
summary of evidence as to thickness of, 56
yields to pressure, 44
Elephant Hawkmoth, disguise of, 189
Environment and discontinuity, 353, 355
effect of change of, 298
may differ in same locality, 356
Erosion of lake-basins, conditions favourable to, 100
of lakes proved by their forms and contours, 113, 120
Erratic blocks and ice-sheets, 59
Erratics and perched blocks, 69
distribution of British, 88
in Great Britain, 80
in North America, 74
in Switzerland, 71
on the Jura, 72
Erskine, Admiral, on Polynesians, 410
European plants not in N. America, 204
Evolution and distribution of animals, 267
the method of organic, 345
Eyes, rudimentary, 326
Eyre, Mr., on character of Australians, 472
F.
Faults absent beneath oceans, 35
Fear not instinctive, 499
Ferguson on Cambodian architecture, 480
Finger-marks and organic stability, 371
not a specific character, 372
Fisher, Rev. O., on earth's crust, 41
on permanence of oceans, 28
Flora of Brazilian Campos, 309
of Britain and Eastern States compared, 201
N. America and Europe, relations of, 200
of Sierra Nevada and California, 223
of the Californian forests, 226
Prof. Asa Gray on, 200
Sir Joseph Hooker on, 201
Floral beauty in England and America, 210
Flowers and forests of the far west, 213
Flowers, English and American, 199
Henslow's theory of modification of, 306
breaksdown, 308
in Michigan in March, 207
„ „ in July, 208
near Cincinnati in April, 207
near Washington in February, 206
rambles after American, 206
Four limbs of vertebrates, 360
Four toes of birds, 360
Forests, destruction of in N.E. America, 199
of California, 224
Forms of lakes prove erosion, 113
G.
Galapagos, few spiny plants in, 309
Galls, a crucial case of non-heredity, 307
Galton, F., misconceives natural selection, 367
on checks to population, 521
on finger-marks, 371
on improvement of the race, 513
on positions of organic stability, 366
theory of, 315
Gardner, J. S., on crust-movements, 46
Garnier on ruins of Baioñ, 478
Gätke on bird-migration, 503
Geikie, Prof. J., on origin of the northern drift, 108
on Lake Zurich as a supposed case of earth movement, 143
Sir A., on thick deposits in shallow water, 44
Genera, discontinuous in S. America, 273
how explained, 274
how they originate, 299
origin and increase of, 268
occasional discontinuity of, 271
Genius appears suddenly, 319
Geological record on Continents, 36
Geometer larvæ, disguise of, 193
Giant tortoises, distribution of, 281
Gibbons, 161
Giles on beauty in Australians, 470
Giraffe, how developed, 329
Glaciers, eroding power of, 107
moving up hill, 75, 80
Gorges, formed by sub-glacial torrents, 119, 120
narrow water-worn, 134
distribution of, 135
Gorilla, 152
Govett's Leap, 12
Granite, forms assumed by, 22
decomposition of, 24
Grant Allen on race-improvement, 514
Gravity, variations of force of, 47
Gray's Peak, alpine flora of, 218
Green river valley, how formed, 17
Grey, Sir G., on Australian rock-paintings, 469
Grimsel Pass, 129
Hospice, 130
Grose valley, 11
Growth, laws of, 375
H.
Habits, supposed inheritance of, 320
Hairy Aino, 467
Hairy Australians, 466
Hanuman, the, 163
Hapalidæ, 174
Heligoland, bird-migration in, 503
Henslow on origin of flowers, 306
Herbivora cause spines in plants, 311
Heredity, experiments on, 316
Hitchcock, Prof., on erratics of Mount Washington, 75
on the Ainos, 475
Homing-instinct, alleged, 505
Horns, origin of, 321
Howling monkeys, 166
Howorth, Sir H., denies possibility of ice-sheets, 77
on Swiss erratics, 73
Human selection, 509
Huxley on utility, 380
Hylobates, 161
I
Ice-age and its work, 59
Ice-eddies in Spitzbergen, 138
Ice-erosion of lakes, criticism of, 143
objections to, 104
of lake-basins, objections to, 120
summary of evidence for, 126
Ice-sheet of the Rhone valley, 78
the N. American, 76
Ice-sheets of British Isles, 81
Inaccessible valleys, formation of, 19
Increase, illustration of rapid, 368
Insects, five parts of legs of, 360
the disguises of, 185
Instinct, the problem of, 497, 501
Iphias glaucippe, a leaf-butterfly, 190
J.
Jaws of civilised man, small, 331
Judd, Prof., on volcanic phenomena, 51
Jukes-Browne, on oceanic islands, 29, 34
K.
Keane, Prof., on primitive Caucasians, 475
on affinities of Mahoris, 413
Kelvin, Lord, on liquid spheroid, 43
Kendal, Mr. P. F., on teachings of British erratics, 88
Kerner, A., on spiny plants and herbivora, 311
Khmers of Cambodia, 475, 481, 484
Kilauea and Mauna Loa, volcanic phenomena of, 54
Kirchet, the, 131
L.
Lagothrix, 168
Lagoa Santa, flora of, 309
Lake-basins, Desor and Falsan's theory of, 124
objections to, 125
explained by earth-movements, 109
glacial erosion of, 94
objections to the theory, 110
Lakes, conditions favourable to ice-erosion of, 100
different kinds of, 95
distribution of, 94
scarce in non-glaciated countries, 98
Lake Como, section of, 103
Lake of Geneva, a Test of Rival Theories, 120
Lamarck resuscitated in America, 347
Lamarckism and Darwinism, 350
Languages of Australia and Africa, 464
Lappet-moth, disguise of, 187
Lava, outflow of how explained, 54
stream cut through by river, 18
Lawes, Mr., on the Motu, 451
Laws of growth, 375
Leaf-butterfly, the Indian, 191
habits of, 193
Leaf-insects, various, 195
Lemurs, 176
distribution of, 280
Lemur, ring-tailed, 179
Leprosy in the Sandwich Is., 408
Lincoln, Dr. D. F., on ice-erosion of Seneca Lake, 141
Locusts at sea, 251
VOL. I. M M
Loris, 176
Lyell, Sir C., gives up iceberg theory to explain Swiss erratics, 73
on sinking of land, 45
on sub-glacial moraine at Soleure, 79
M.
Macacus, 164
Madagascar and West Indies, 278
lemurs of, 179
Madeira, coleoptera of, 250
Mahoris are of Caucasian affinities, 412
or brown Polynesians, 404
Malays and Polynesians compared, 402
Mammals, cervical vertebræ of, 361
of New Guinea, 419
Man, jaws of civilised small, 331
Mantidæ, disguises of, 197
Map of Ancient Rhone Glacier, explanation of, 128
Marmosets, 174
Mentawi islands, natives of, 490
Merriam, Mr. C. H., on migration, 503
Mias, 157
nest-building of, 159
Midas chrysoleucas, 175
Migratory instinct, 502
Migration, report of Committee of B.A. on, 505
Miklucho Maclay, Dr., on Papuans, 454
Mississippi valley, migration in, 503
Mivart, Dr., on recognition marks, 384
Modification in the individual, 338
Moel Tryfaen, glacial gravels of, 92
Monkeys, affinities and distribution of, 146
affinities of, 182
African, 152
American, 165
Asiatic, 157
difference from men, 148
different kinds of, 149
distribution of, 180
feet and hands of, 147
geological range of, 181
Old World and American, 150
Monstrosities and origin of species, 358
Moraines, 62
Moraine at Flamborough Head, 83
at Thornwick on chalk, 83
Morgan, C. Lloyd, on instinct, 497
on inheritance of acquired characters, 507
Moselle river illustrating valley formation, 17
Moths resembling lichen, 197
Motu tribe in N. Guinea, 450
Mouhot on old Cambodian temples, 476
Mounds in N. W. Australia, 470
Mountains, roots of, 47
Moura, J., portraits of Cambodians, 481
Murray, Andrew, on distribution of beetles, 250
Sir J., on permanence of oceans, 28
on depth of oceans, 30
Mycetes, 166
N.
Natives of New Hebrides, 457
of Timor, 458
Natural selection will improve our race 523
Nepean river, N.S. Wales, 20
New Guinea, 416
birds of, 423, 433
climate of, 418
early visitors to, 417
inhabitants of, 434
mammals of, 419
New species, how originated, 290
Niata cattle, 393
Night-monkeys, 174
N. America, botanical divisions of, 213
rural scenery of, 209
weeds are mostly European, 210
N. American plants not in Europe, 205
North Australia, native of, 493
Number of parts, variation in, 359
Nuts, not a direct adaptation, 312
Nycticebus, 176
Nyctipithecus, 174
O.
Ocean floor, contour of, 34
Ocean floor and land surface compared, 35
Oceanic and continental masses, 31
Oceans not raised and depressed, 35
Onochycerus scorpio, disguise of, 194
Orange tip butterfly, disguise of, 189
Orang-utan, 157
Organic stability, 366
Origin of species and genera, 285
P
Palæarctic and Nearctic regions, 235
land-birds of, 241
mammals of, 237
summary of facts, 247
Papuans, 434
affinities of, 459
characters of, 436
customs of, 446
hair of, 438
houses of, 440
manufactures of, 442
Paradise-birds, 423
new species of, 426
number of, 432
Pendulum experiments, 47
Perched blocks, 69, 86
Permanence of ocean basins, 28, 37
Pines of the Sierra Nevada, 225
Pipits, meadow and tree, 356
Pithecia, 171
Pitta angolensis, isolation of, 273
Plains and prairies, the, 214
Platt Ball on use and disuse, 335
Polynesian languages, 401
Polynesians, affinities of, 461, 496
Polynesians and Malays compared, 402
affinities of, 404
Darwin, on character of, 405
Polynesians and their migrations, 399
origin of brown, 489
tribes in N. Guinea, 450
Population, checks to increase of, 520
Porto Santo, rabbits of, 393
Prestwich, Prof., on ocean depths, 30
on heat of earth, 42
Prickles, origin of, 308
Proteus, rudimentary eyes of, 326
white colour of, 393
Q.
Quatrefages on Polynesians, 399
their migrations, 400
Queensland, natives of, 462
R.
Rabbits of Porto Santo, 393
Recognition-marks, 383
criticism of, 384
Redwood, 232
Report on bird-migration, 505
Rhone, ancient glacier of, 78
Roches moutonnées, 64
Rocks, striated, grooved, and fluted, 67
Rocky mountain flora, 215
Rocky mountains, British plants in 216, 219
alpine plants of, 217
origin of alpine flora of, 221
Rock-paintings in Australia, 469
Romanes on scent in dogs, 506
S.
Sakis, 171
Salt rocks favour denudation, 20
Samoa, natives of, 412
Sandstone, caverns in, 20
Sandwich Islands, few spiny plants in, 309
natives of, 406
Sapajous, 170
Sarcodes sanguinea, 226
Saturnia, Texan, change in, 338
Scandinavian boulders in Yorkshire, 86
rocks in Britain, 82
Scrope on volcanic phenomena, 51
Sedgwick, Prof., on Scandinavian erratics in Britain, 82
Seebohm on migration, 504
Selection, human, 509
of qualities not characters, 332
Semnopithecus, 163
Semon on Australians and Caucasians, 468
Sense of direction, no proof of, 506
Sequoia sempervirens, 232
Sequoias, age of, 230
few young trees, 231
Shap granite, erratics of, 88
Sierra Nevada, flora of, 223
Simia satyrus, 160
Social advance and character, 517
Spalding, D., on instinct, 498
Species, how they originate, 290
meaning of, 285
why stable, 381
Specific characters due to natural selection, 385
must therefore be useful, 387
supposed useless, 393
Spectre-lemur, 177
Spencer, H., erroneous assumptions of, 330
on the population question, 522
on touch, 324
Spencer, Prof., on Central Australians, 475
photographs of Australians, 490
Spider monkeys, 167
Spines, origin of, 308
Spiny plants and herbivorous animals, 310
rare in oceanic islands, 309
abundant in Africa and Chile, 310
frequent in moist climates, 310
Stability of species, 381
organic, 366, 369
Stanton, Eliz. C., on true marriage, 515
Stanley, Hiram M., on improvement of the race, 513
Stick-insects, 195
Stone, Mr. O. C., on the Motu tribe, 450
Striated rocks, 67
Sugar-pines, 227
T.
Tahiti, natives of, 405
Tarr, Mr. R. S., on Lake Cayuga, an ice-eroded basin, 141
Tarsius, 177
Teeth, evolution of, 323
Temperature underground, variations of under mountains, 47
Thomson, Mr. J., on ruins in Cambodia, 477
Tiger beetles, disguises of, 194
Tonga Is., natives of, 410
Tortoise-shell butterfly, chrysalis of, 197
Training, are results of hereditary, 510
Trientalis europæa in California, 227
Tropical regions, allied forms in, 275
explanation of, 276, 278
Turner, Rev. W., on the Motu, 452
U.
Uacari monkeys, 171
Upward motion of rocks in glaciers of Spitzbergen, 91
Use-inheritance not a factor in evolution, 342
Utility of specific characters, 364
a result of natural selection, 387
conclusions as to, 397
Romanes on, 378
special difficulties, 393
the principle of, 379
the problem of, 378
V
Valleys, how formed, 14
Variability, 292
of all observed characters, 297
of many parts at once, 294
Variation, cause of unknown, 302
an ultimate fact in nature, 302
definite and indefinite, 305
Variations, discontinuous, 357
Varieties, intrinsic and extrinsic, 314
Veddahs of Ceylon, 486
Venetz, on former extension of Swiss glaciers, 61
Vertebrates, four limbs of, 360
Volcanoes, as disproving liquid earth, 43
Volcanic mountains have no roots, 49
Volcanic phenomena, how explained, 50
illustrated by molten silver, 52
W.
Walking-leaf, the Indian, 185
Warming, E., on spiny plants in Brazil, 309
Weismann, theory of, 315
West Indies and Madagascar, 278
Whitney, on Yosemite, 7, 25
Wollaston, on beetles of Madeira, 252
Woolly monkeys, 168
Wright, Prof. G. F., on N. American erratics, 74
Y.
Yosemite valley, 4, 21
supposed origin of, 7
Z.
Zool. distribution principles of, 282
Zurich, Lake, not due to earth-movements, 144
RICHARD CLAY AND SONS, LIMITED, LONDON AND BUNGAY.